

Alternative Carbon Sources for Isoprene Emission
- PMID: 30472998
- PMCID: PMC6354897
- DOI: 10.1016/j.tplants.2018.09.012
Alternative Carbon Sources for Isoprene Emission
Abstract
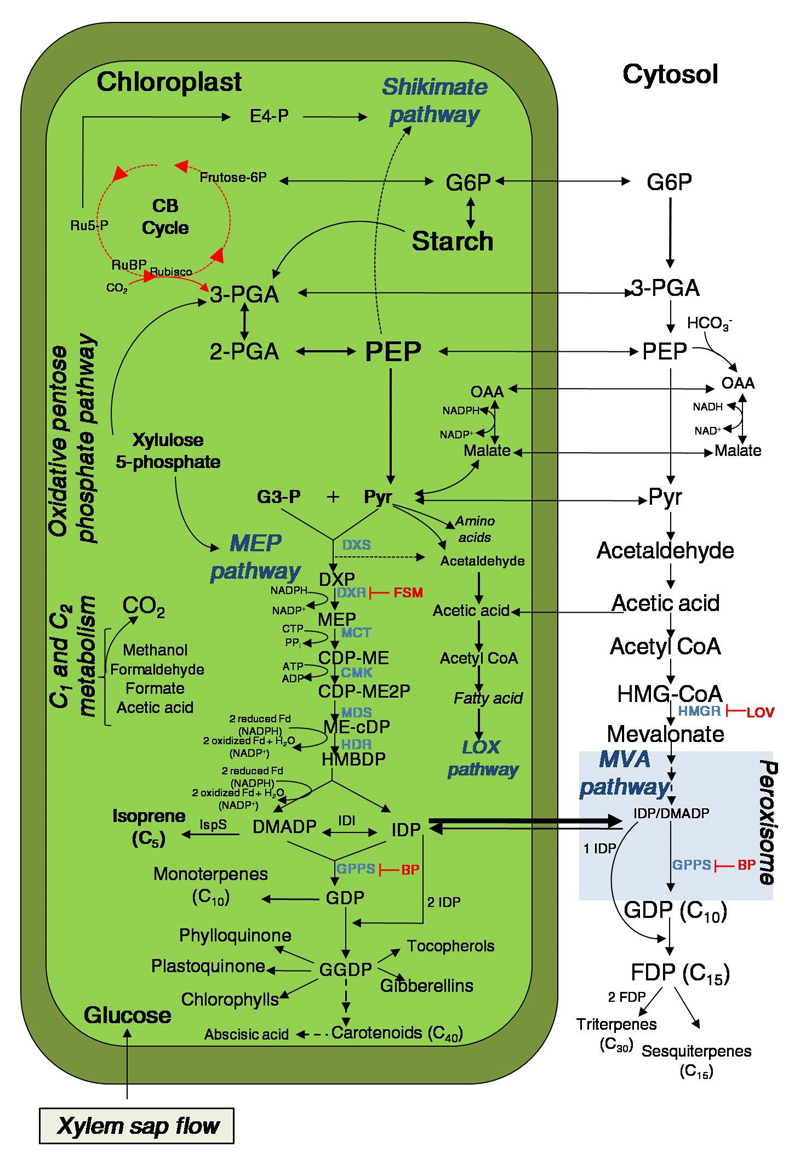
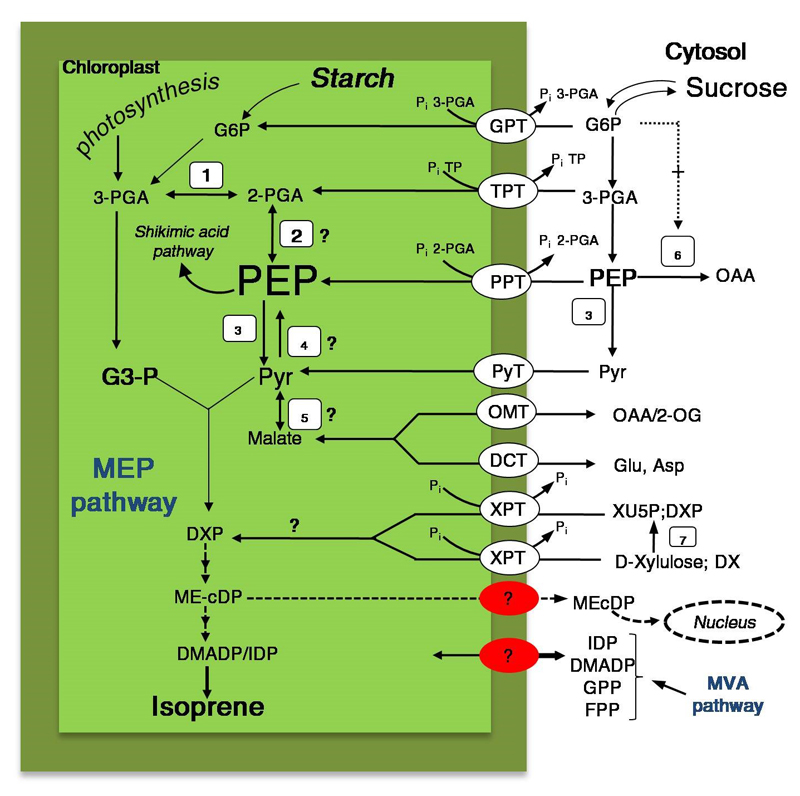
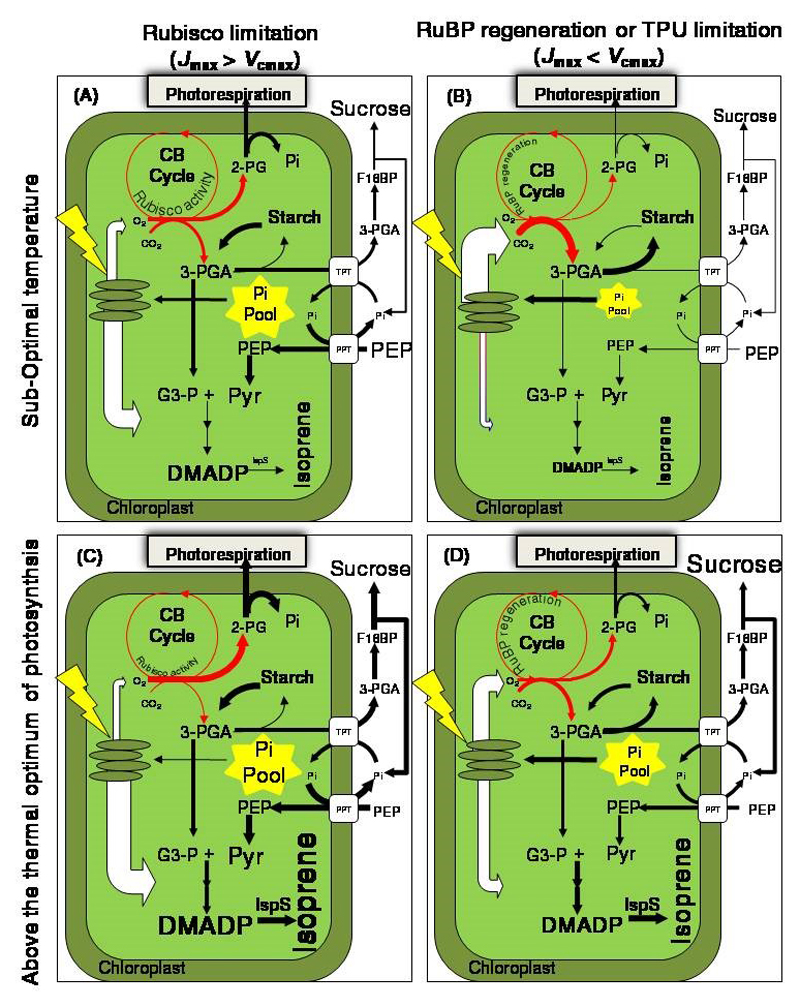
Isoprene and other plastidial isoprenoids are produced primarily from recently assimilated photosynthates via the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway. However, when environmental conditions limit photosynthesis, a fraction of carbon for MEP pathway can come from extrachloroplastic sources. The flow of extrachloroplastic carbon depends on the species and on leaf developmental and environmental conditions. The exchange of common phosphorylated intermediates between the MEP pathway and other metabolic pathways can occur via plastidic phosphate translocators. C1 and C2 carbon intermediates can contribute to chloroplastic metabolism, including photosynthesis and isoprenoid synthesis. Integration of these metabolic processes provide an example of metabolic flexibility, and results in the synthesis of primary metabolites for plant growth and secondary metabolites for plant defense, allowing effective use of environmental resources under multiple stresses.
Keywords: DMADP; MEP pathway; VOCs; energy control; plant stress.
Copyright © 2018. Published by Elsevier Ltd.
Conflict of interest statement
The authors declare that there are no conflicts of interest
Figures
References
-
- Kessler A. Introduction to a Virtual Special Issue on plant volatiles. New Phytol. 2016;209:1333–1337. - PubMed
-
- Dudareva N, et al. Plant Volatiles : Recent Advances and Future Perspectives. Landscape. 2006 doi: 10.1080/07352680600899973. - DOI
-
- Guenther AB, et al. The model of emissions of gases and aerosols from nature version 2.1 (MEGAN2.1): An extended and updated framework for modeling biogenic emissions. Geosci Model Dev. 2012;5:1471–1492.
-
- Henrot A-J, et al. Implementation of the MEGAN (v2.1) biogenic emission model in the ECHAM6-HAMMOZ chemistry climate model. Geosci Model Dev. 2017;10:903–926.
-
- Unger N. Isoprene emission variability through the twentieth century. J Geophys Res Atmos. 2013;118:13,606–13,613.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
