

NuA4 acetyltransferase is required for efficient nucleotide excision repair in yeast
- PMID: 30473425
- PMCID: PMC6312484
- DOI: 10.1016/j.dnarep.2018.11.006
NuA4 acetyltransferase is required for efficient nucleotide excision repair in yeast
Abstract
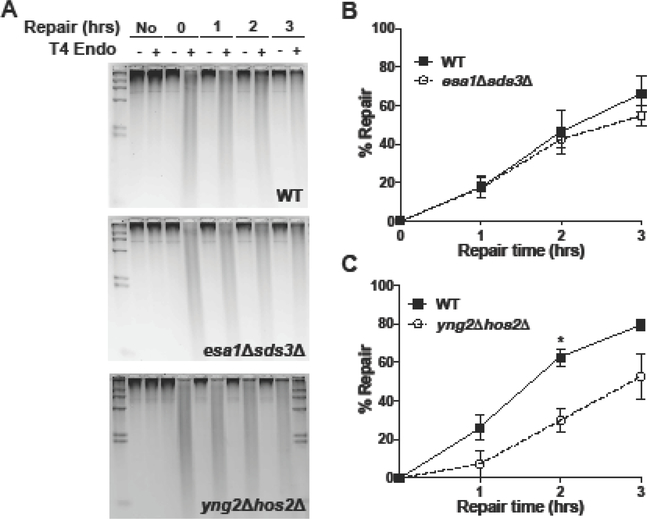
The nucleotide excision repair (NER) pathway is critical for removing damage induced by ultraviolet (UV) light and other helix-distorting lesions from cellular DNA. While efficient NER is critical to avoid cell death and mutagenesis, NER activity is inhibited in chromatin due to the association of lesion-containing DNA with histone proteins. Histone acetylation has emerged as an important mechanism for facilitating NER in chromatin, particularly acetylation catalyzed by the Spt-Ada-Gcn5 acetyltransferase (SAGA); however, it is not known if other histone acetyltransferases (HATs) promote NER activity in chromatin. Here, we report that the essential Nucleosome Acetyltransferase of histone H4 (NuA4) complex is required for efficient NER in Saccharomyces cerevisiae. Deletion of the non-essential Yng2 subunit of the NuA4 complex causes a general defect in repair of UV-induced cyclobutane pyrimidine dimers (CPDs) in yeast; in contrast, deletion of the Sas3 catalytic subunit of the NuA3 complex does not affect repair. Rapid depletion of the essential NuA4 catalytic subunit Esa1 using the anchor-away method also causes a defect in NER, particularly at the heterochromatic HML locus. We show that disrupting the Sds3 subunit of the Rpd3L histone deacetylase (HDAC) complex rescued the repair defect associated with loss of Esa1 activity, suggesting that NuA4-catalyzed acetylation is important for efficient NER in heterochromatin.
Keywords: Acetylation; Chromatin; DNA repair; Esa1; Histone; NER.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures





Similar articles
-
Using Yeast as a Model Organism to Study the Functional Roles of Histone Acetylation in DNA Excision Repair.Methods Mol Biol. 2019;1983:175-190. doi: 10.1007/978-1-4939-9434-2_10. Methods Mol Biol. 2019. PMID: 31087298 Free PMC article.
-
The Saccharomyces cerevisiae histone acetyltransferase Gcn5 has a role in the photoreactivation and nucleotide excision repair of UV-induced cyclobutane pyrimidine dimers in the MFA2 gene.J Mol Biol. 2002 Feb 22;316(3):489-99. doi: 10.1006/jmbi.2001.5383. J Mol Biol. 2002. PMID: 11866513
-
Histone variant Htz1 promotes histone H3 acetylation to enhance nucleotide excision repair in Htz1 nucleosomes.Nucleic Acids Res. 2013 Oct;41(19):9006-19. doi: 10.1093/nar/gkt688. Epub 2013 Aug 7. Nucleic Acids Res. 2013. PMID: 23925126 Free PMC article.
-
Histone modification and chromatin remodeling during NER.DNA Repair (Amst). 2015 Dec;36:105-113. doi: 10.1016/j.dnarep.2015.09.013. Epub 2015 Sep 16. DNA Repair (Amst). 2015. PMID: 26422133 Review.
-
Nucleotide excision repair in cellular chromatin: studies with yeast from nucleotide to gene to genome.Int J Mol Sci. 2012;13(9):11141-11164. doi: 10.3390/ijms130911141. Epub 2012 Sep 7. Int J Mol Sci. 2012. PMID: 23109843 Free PMC article. Review.
Cited by
-
Genome-wide maps of UV damage repair and mutation suppression by CPD photolyase.Nucleic Acids Res. 2025 Jun 6;53(11):gkaf495. doi: 10.1093/nar/gkaf495. Nucleic Acids Res. 2025. PMID: 40498072 Free PMC article.
-
Formation and Recognition of UV-Induced DNA Damage within Genome Complexity.Int J Mol Sci. 2020 Sep 12;21(18):6689. doi: 10.3390/ijms21186689. Int J Mol Sci. 2020. PMID: 32932704 Free PMC article. Review.
-
Insights Into the Function of the NuA4 Complex in Plants.Front Plant Sci. 2020 Feb 21;11:125. doi: 10.3389/fpls.2020.00125. eCollection 2020. Front Plant Sci. 2020. PMID: 32153620 Free PMC article. Review.
-
Genome-wide maps of rare and atypical UV photoproducts reveal distinct patterns of damage formation and mutagenesis in yeast chromatin.Proc Natl Acad Sci U S A. 2023 Mar 7;120(10):e2216907120. doi: 10.1073/pnas.2216907120. Epub 2023 Feb 28. Proc Natl Acad Sci U S A. 2023. PMID: 36853943 Free PMC article.
-
Variable impact of conformationally distinct DNA lesions on nucleosome structure and dynamics: Implications for nucleotide excision repair.DNA Repair (Amst). 2020 Mar;87:102768. doi: 10.1016/j.dnarep.2019.102768. Epub 2019 Dec 28. DNA Repair (Amst). 2020. PMID: 32018112 Free PMC article.
References
-
- Friedberg EC, Walker GC, Siede W, Wood RD, Schultz RA, Ellenberger T, DNA repair and Mutagenesis, 2nd ed., ASM Press, Washington, D.C., 2006.
-
- Marteijn JA, Lans H, Vermeulen W, Hoeijmakers JH, Understanding nucleotide excision repair and its roles in cancer and ageing, Nat Rev Mol Cell Biol, 15 (2014) 465–481. - PubMed
-
- Hanawalt PC, Spivak G, Transcription-coupled DNA repair: two decades of progress and surprises, Nat Rev Mol Cell Biol, 9 (2008) 958–970. - PubMed
-
- Li S, Transcription coupled nucleotide excision repair in the yeast Saccharomyces cerevisiae: The ambiguous role of Rad26, DNA repair, 36 (2015) 43–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
