Extracellular membrane vesicles in the three domains of life and beyond
- PMID: 30476045
- PMCID: PMC6524685
- DOI: 10.1093/femsre/fuy042
Extracellular membrane vesicles in the three domains of life and beyond
Abstract
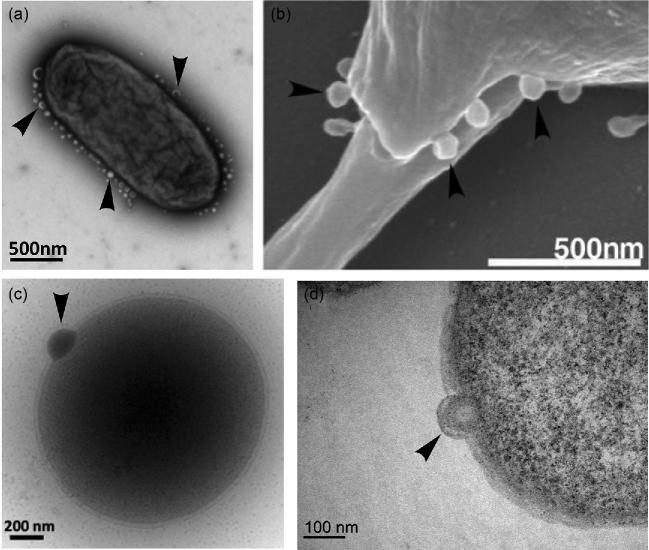
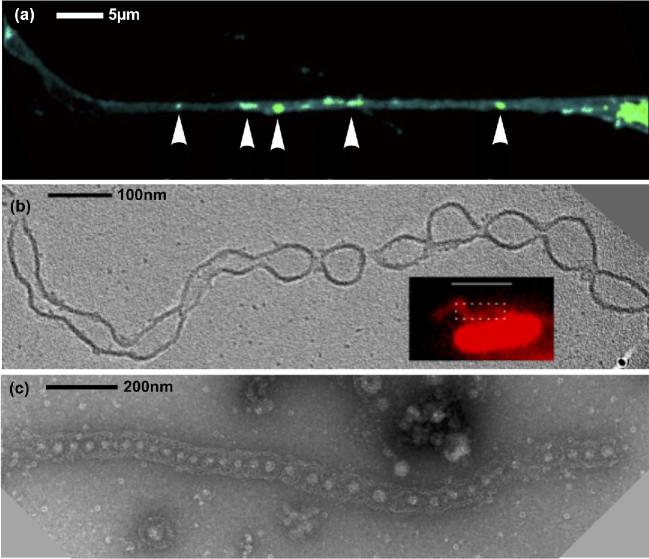
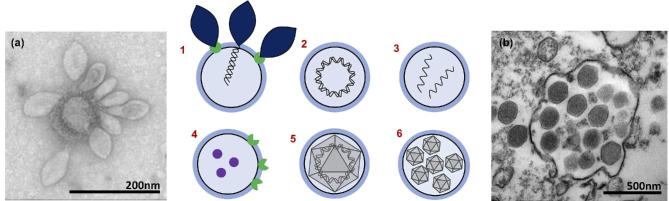
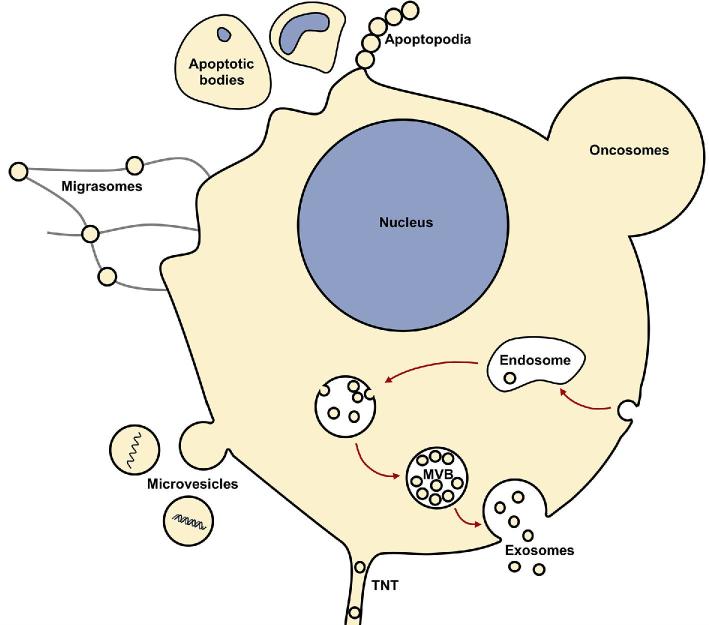
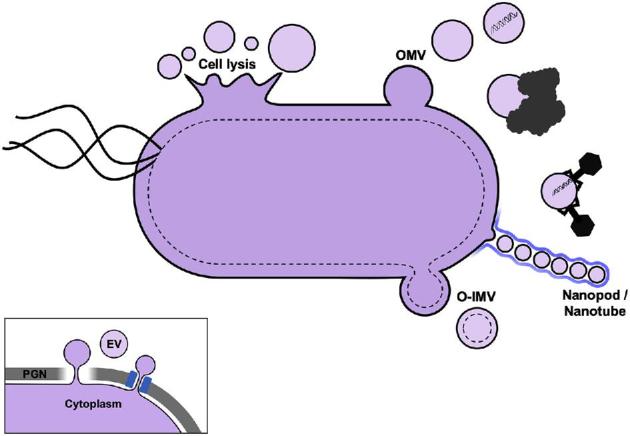
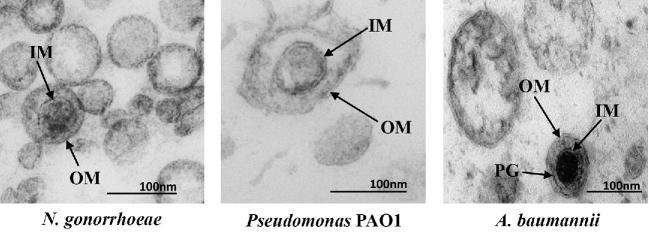
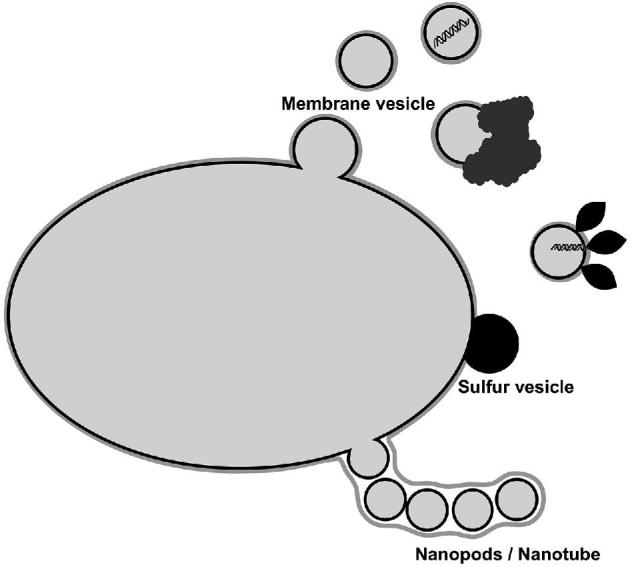
Cells from all three domains of life, Archaea, Bacteria and Eukarya, produce extracellular vesicles (EVs) which are sometimes associated with filamentous structures known as nanopods or nanotubes. The mechanisms of EV biogenesis in the three domains remain poorly understood, although studies in Bacteria and Eukarya indicate that the regulation of lipid composition plays a major role in initiating membrane curvature. EVs are increasingly recognized as important mediators of intercellular communication via transfer of a wide variety of molecular cargoes. They have been implicated in many aspects of cell physiology such as stress response, intercellular competition, lateral gene transfer (via RNA or DNA), pathogenicity and detoxification. Their role in various human pathologies and aging has aroused much interest in recent years. EVs can be used as decoys against viral attack but virus-infected cells also produce EVs that boost viral infection. Here, we review current knowledge on EVs in the three domains of life and their interactions with the viral world.
Keywords: Archaea; LUCA; evolution; extracellular vesicles; nanotubes; virus.
© FEMS 2018.
Figures