

Overexpression of thioredoxin m in tobacco chloroplasts inhibits the protein kinase STN7 and alters photosynthetic performance
- PMID: 30476130
- PMCID: PMC6363096
- DOI: 10.1093/jxb/ery415
Overexpression of thioredoxin m in tobacco chloroplasts inhibits the protein kinase STN7 and alters photosynthetic performance
Abstract
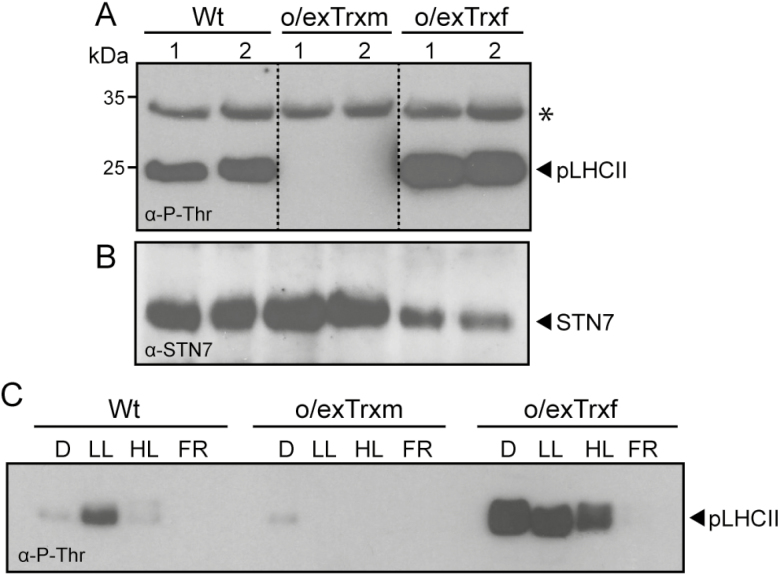
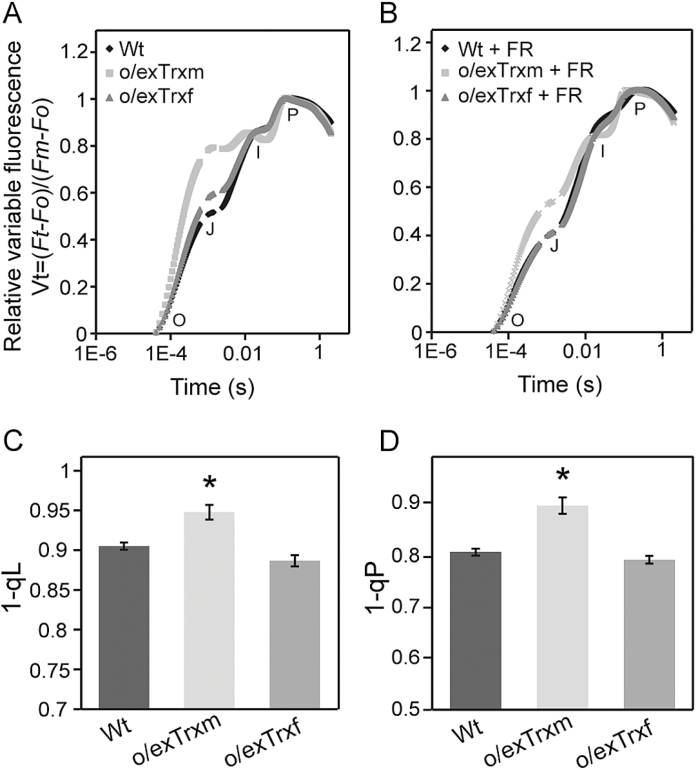
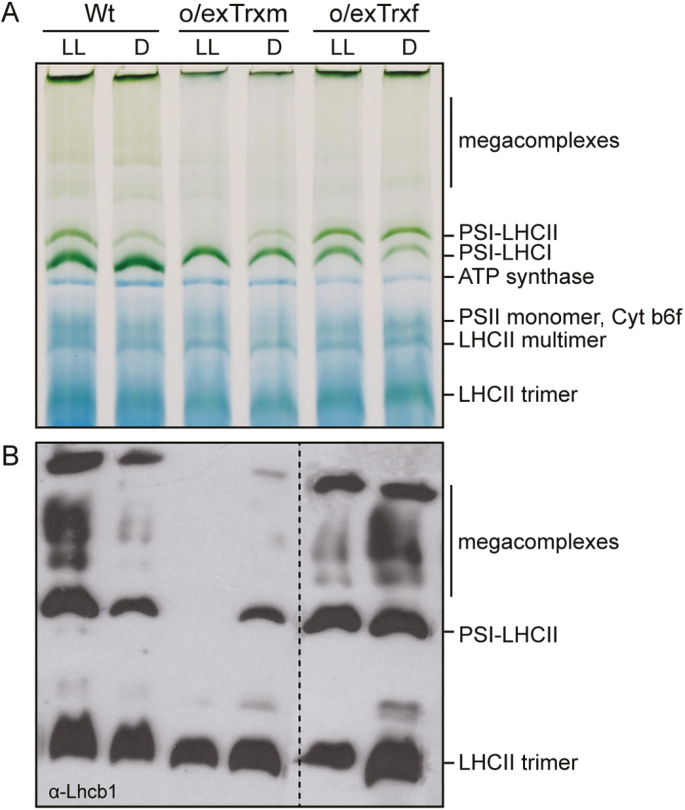
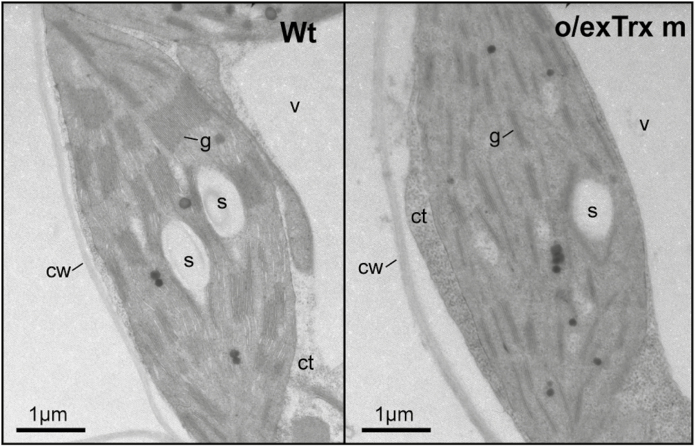
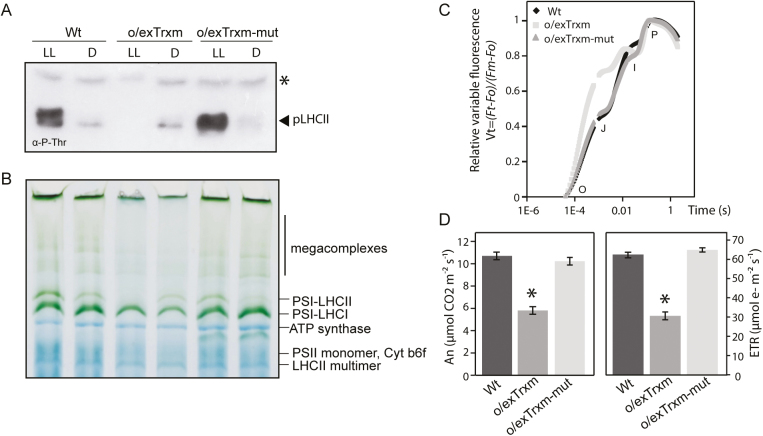
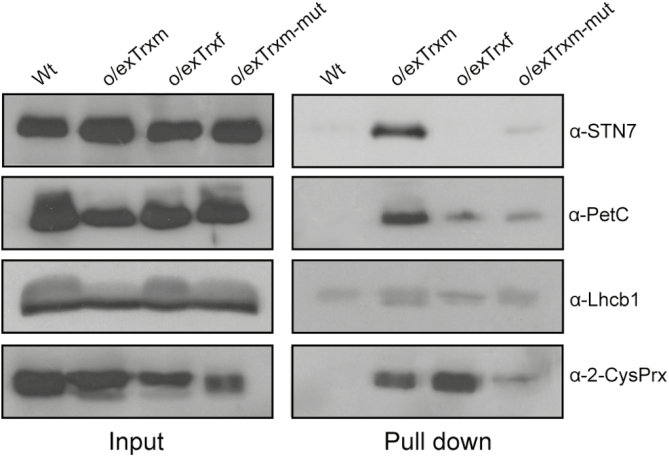
The activity of the protein kinase STN7, involved in phosphorylation of the light-harvesting complex II (LHCII) proteins, has been reported as being co-operatively regulated by the redox state of the plastoquinone pool and the ferredoxin-thioredoxin (Trx) system. The present study aims to investigate the role of plastid Trxs in STN7 regulation and their impact on photosynthesis. For this purpose, tobacco plants overexpressing Trx f or m from the plastid genome were characterized, demonstrating that only Trx m overexpression was associated with a complete loss of LHCII phosphorylation that did not correlate with decreased STN7 levels. The absence of phosphorylation in Trx m-overexpressing plants impeded migration of LHCII from PSII to PSI, with the concomitant loss of PSI-LHCII complex formation. Consequently, the thylakoid ultrastructure was altered, showing reduced grana stacking. Moreover, the electron transport rate was negatively affected, showing an impact on energy-demanding processes such as the Rubisco maximum carboxylation capacity and ribulose 1,5-bisphosphate regeneration rate values, which caused a strong depletion in net photosynthetic rates. Finally, tobacco plants overexpressing a Trx m mutant lacking the reactive redox site showed equivalent physiological performance to the wild type, indicating that the overexpressed Trx m deactivates STN7 in a redox-dependent way.
Figures
References
-
- Allen JF. 1992. Protein phosphorylation in regulation of photosynthesis. Biochimica et Biophysica Acta 1098, 275–335. - PubMed
-
- Allen JF. 2003. Botany. State transitions—a question of balance. Science 299, 1530–1532. - PubMed
-
- Allen JF. 2017. Why we need to know the structure of phosphorylated chloroplast light-harvesting complex II. Physiologia Plantarum 161, 28–44. - PubMed
-
- Belkhodja R, Morales F, Quilez R, López-Millán AF, Abadía A, Abadía J. 1998. Iron deficiency causes changes in chlorophyll fluorescence due to the reduction in the dark of the Photosystem II acceptor side. Photosynthesis Research 56, 265–276.
-
- Bellafiore S, Barneche F, Peltier G, Rochaix JD. 2005. State transitions and light adaptation require chloroplast thylakoid protein kinase STN7. Nature 433, 892–895. - PubMed
