

Structural gymnastics of RAG-mediated DNA cleavage in V(D)J recombination
- PMID: 30476719
- PMCID: PMC6457668
- DOI: 10.1016/j.sbi.2018.11.001
Structural gymnastics of RAG-mediated DNA cleavage in V(D)J recombination
Abstract
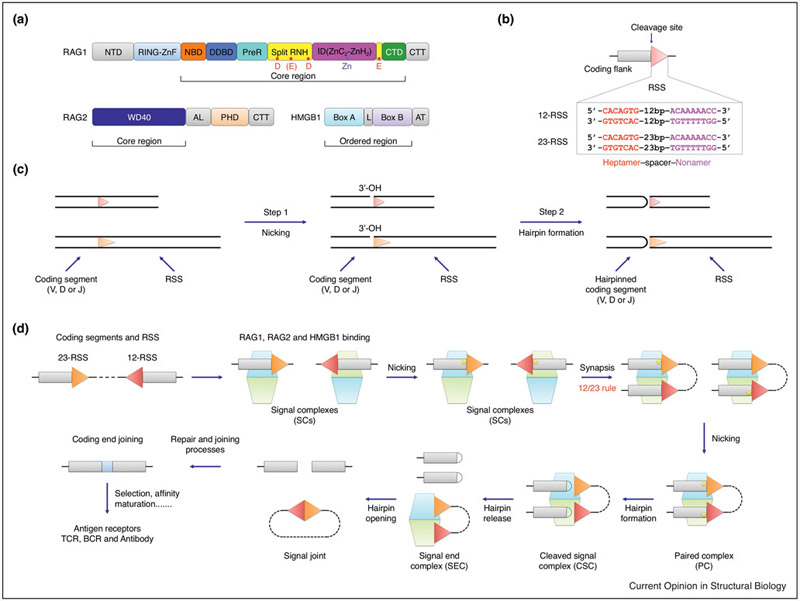
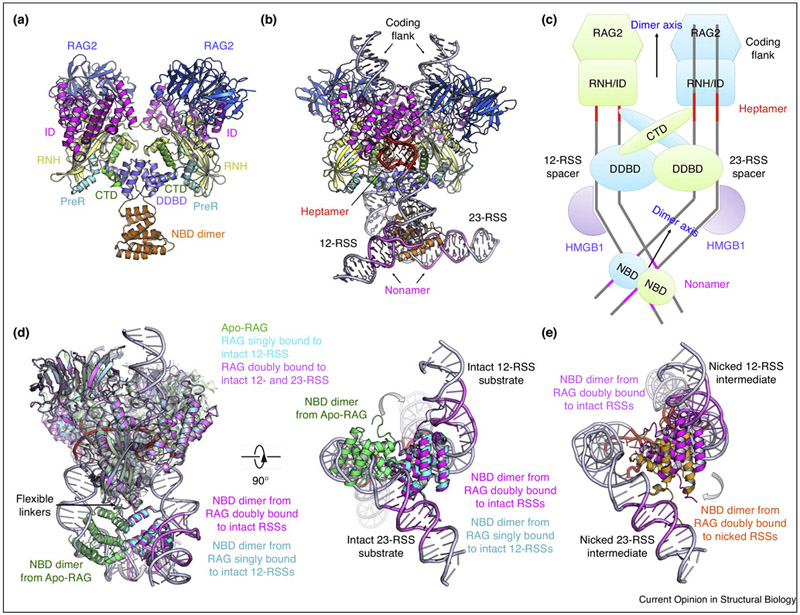
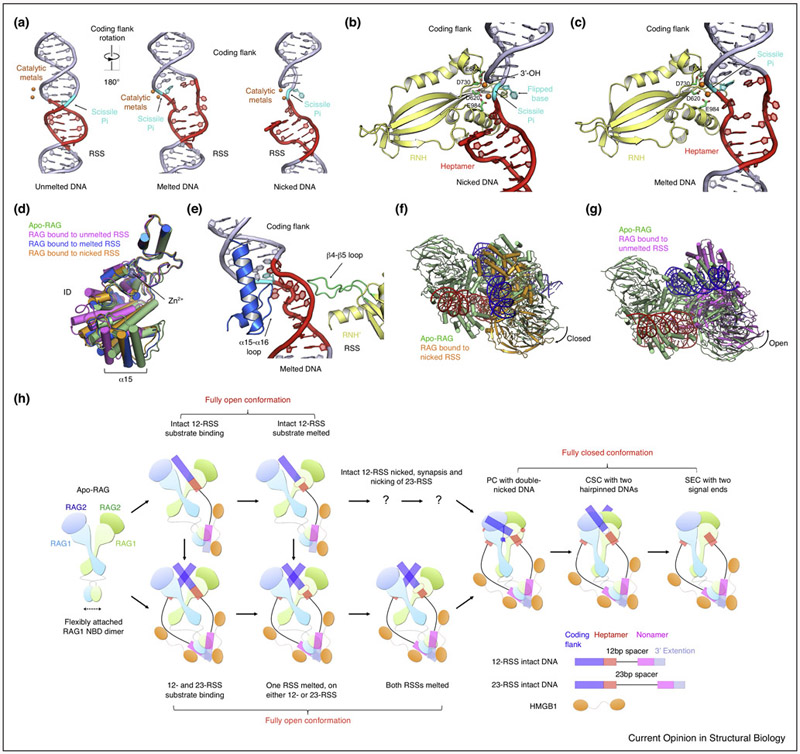
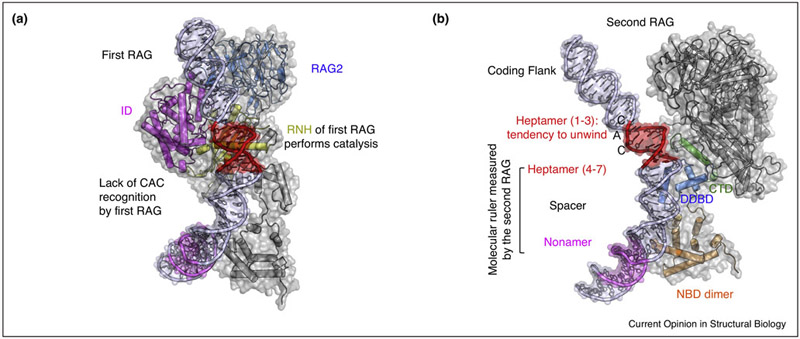
A hallmark of vertebrate immunity is the diverse repertoire of antigen-receptor genes that results from combinatorial splicing of gene coding segments by V(D)J recombination. The (RAG1-RAG2)2 endonuclease complex (RAG) specifically recognizes and cleaves a pair of recombination signal sequences (RSSs), 12-RSS and 23-RSS, via the catalytic steps of nicking and hairpin formation. Both RSSs immediately flank the coding end segments and are composed of a conserved heptamer, a conserved nonamer, and a non-conserved spacer of either 12 base pairs (bp) or 23 bp in between. A single RAG complex only synapses a 12-RSS and a 23-RSS, which was denoted the 12/23 rule, a dogma that ensures recombination between V, D and J segments, but not within the same type of segments. This review recapitulates current structural studies to highlight the conformational transformations in both the RAG complex and the RSS during the consecutive steps of catalysis. The emerging structural mechanism emphasizes distortion of intact RSS and nicked RSS exerted by a piston-like motion in RAG1 and by dimer closure, respectively. Bipartite recognition of heptamer and nonamer, flexibly linked nonamer-binding domain dimer relatively to the heptamer recognition region dimer, and RSS plasticity and bending by HMGB1 together contribute to the molecular basis of the 12/23 rule in the RAG molecular machine.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
There are no conflicts of interest to declare.
Figures
Similar articles
-
Molecular Mechanism of V(D)J Recombination from Synaptic RAG1-RAG2 Complex Structures.Cell. 2015 Nov 19;163(5):1138-1152. doi: 10.1016/j.cell.2015.10.055. Epub 2015 Nov 5. Cell. 2015. PMID: 26548953 Free PMC article.
-
Synergistic requirement of orphan nonamer-like elements and DNA bending enhanced by HMGB1 for RAG-mediated nicking at cryptic 12-RSS but not authentic 12-RSS.Genes Cells. 2011 Aug;16(8):879-95. doi: 10.1111/j.1365-2443.2011.01534.x. Epub 2011 Jul 11. Genes Cells. 2011. PMID: 21740486
-
HMGB1/2 can target DNA for illegitimate cleavage by the RAG1/2 complex.BMC Mol Biol. 2009 Mar 24;10:24. doi: 10.1186/1471-2199-10-24. BMC Mol Biol. 2009. PMID: 19317908 Free PMC article.
-
V(D)J recombination: mechanisms of initiation.Annu Rev Genet. 2011;45:167-202. doi: 10.1146/annurev-genet-110410-132552. Epub 2011 Aug 19. Annu Rev Genet. 2011. PMID: 21854230 Review.
-
New insights into the evolutionary origins of the recombination-activating gene proteins and V(D)J recombination.FEBS J. 2017 Jun;284(11):1590-1605. doi: 10.1111/febs.13990. Epub 2017 Jan 6. FEBS J. 2017. PMID: 27973733 Free PMC article. Review.
Cited by
-
Building a neural network model to define DNA sequence specificity in V(D)J recombination.Nucleic Acids Res. 2025 Jun 20;53(12):gkaf551. doi: 10.1093/nar/gkaf551. Nucleic Acids Res. 2025. PMID: 40548941 Free PMC article.
-
RAG2 abolishes RAG1 aggregation to facilitate V(D)J recombination.Cell Rep. 2021 Oct 12;37(2):109824. doi: 10.1016/j.celrep.2021.109824. Cell Rep. 2021. PMID: 34644584 Free PMC article.
-
Cutting antiparallel DNA strands in a single active site.Nat Struct Mol Biol. 2020 Feb;27(2):119-126. doi: 10.1038/s41594-019-0363-2. Epub 2020 Feb 3. Nat Struct Mol Biol. 2020. PMID: 32015552 Free PMC article.
-
Structural insights into the evolution of the RAG recombinase.Nat Rev Immunol. 2022 Jun;22(6):353-370. doi: 10.1038/s41577-021-00628-6. Epub 2021 Oct 21. Nat Rev Immunol. 2022. PMID: 34675378 Review.
-
The molecular basis and disease relevance of non-homologous DNA end joining.Nat Rev Mol Cell Biol. 2020 Dec;21(12):765-781. doi: 10.1038/s41580-020-00297-8. Epub 2020 Oct 19. Nat Rev Mol Cell Biol. 2020. PMID: 33077885 Free PMC article. Review.
References
-
- Fanning L, Connor A, Baetz K, Ramsden D, Wu GE: Mouse RSS spacer sequences affect the rate of V(D)J recombination. Immunogenetics 1996, 44:146–150. - PubMed
-
- Tonegawa S: Somatic generation of antibody diversity. Nature 1983, 302:575–581. - PubMed
-
- Early P, Huang H, Davis M, Calame K, Hood L: An immunoglobulin heavy chain variable region gene is generated from three segments of DNA: VH, D and JH. Cell 1980, 19:981–992. - PubMed
-
- Taccioli GE, Rathbun G, Oltz E, Stamato T, Jeggo PA, Alt FW: Impairment of V(D)J recombination in double-strand break repair mutants. Science 1993, 260:207–210. - PubMed
-
- Sakano H, Maki R, Kurosawa Y, Roeder W, Tonegawa S: Two types of somatic recombination are necessary for the generation of complete immunoglobulin heavy-chain genes. Nature 1980, 286:676–683. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
