

The Core and Holoenzyme Forms of RNA Polymerase from Mycobacterium smegmatis
- PMID: 30478083
- PMCID: PMC6351736
- DOI: 10.1128/JB.00583-18
The Core and Holoenzyme Forms of RNA Polymerase from Mycobacterium smegmatis
Abstract
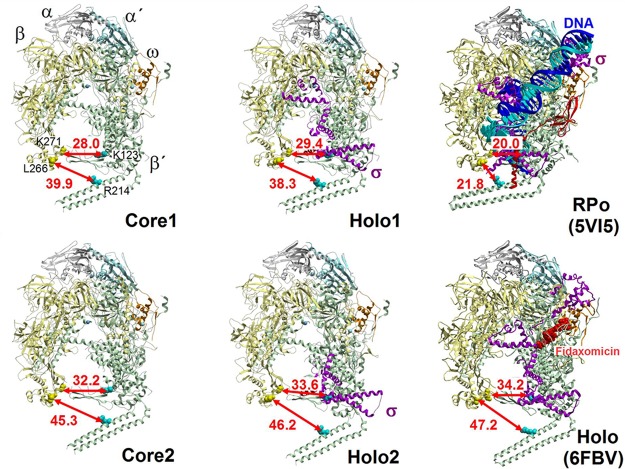
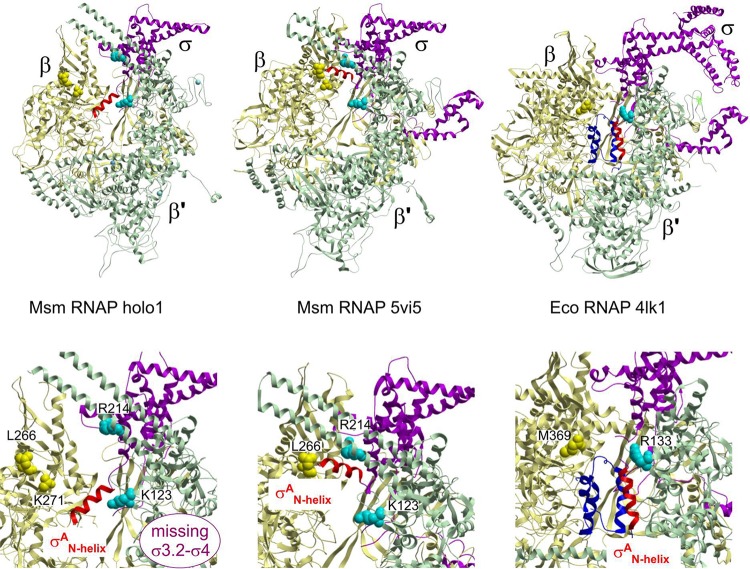
Bacterial RNA polymerase (RNAP) is essential for gene expression and as such is a valid drug target. Hence, it is imperative to know its structure and dynamics. Here, we present two as-yet-unreported forms of Mycobacterium smegmatis RNAP: core and holoenzyme containing σA but no other factors. Each form was detected by cryo-electron microscopy in two major conformations. Comparisons of these structures with known structures of other RNAPs reveal a high degree of conformational flexibility of the mycobacterial enzyme and confirm that region 1.1 of σA is directed into the primary channel of RNAP. Taken together, we describe the conformational changes of unrestrained mycobacterial RNAP.IMPORTANCE We describe here three-dimensional structures of core and holoenzyme forms of mycobacterial RNA polymerase (RNAP) solved by cryo-electron microscopy. These structures fill the thus-far-empty spots in the gallery of the pivotal forms of mycobacterial RNAP and illuminate the extent of conformational dynamics of this enzyme. The presented findings may facilitate future designs of antimycobacterial drugs targeting RNAP.
Keywords: RNA polymerase; bacterial transcription; conformational change; cryo-electron microscopy; mycobacteria; protein structure; transcription initiation factor.
Copyright © 2019 American Society for Microbiology.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
