

Direct Interaction between the Two Z Ring Membrane Anchors FtsA and ZipA
- PMID: 30478085
- PMCID: PMC6351747
- DOI: 10.1128/JB.00579-18
Direct Interaction between the Two Z Ring Membrane Anchors FtsA and ZipA
Abstract
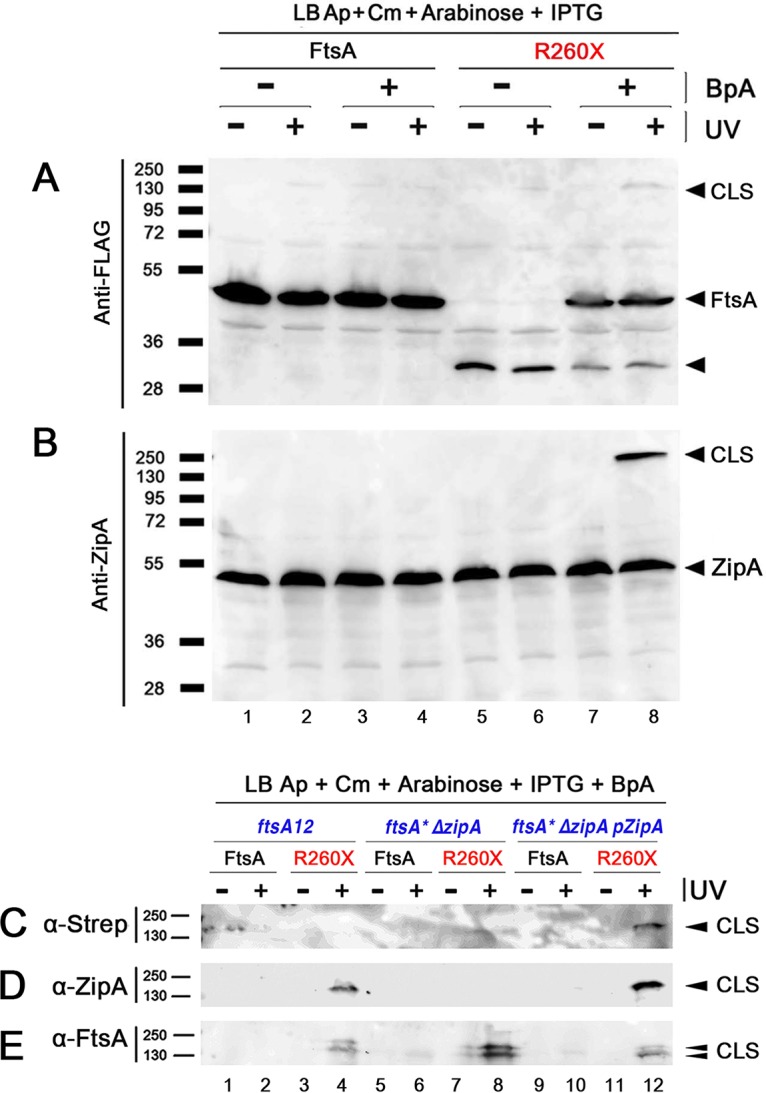
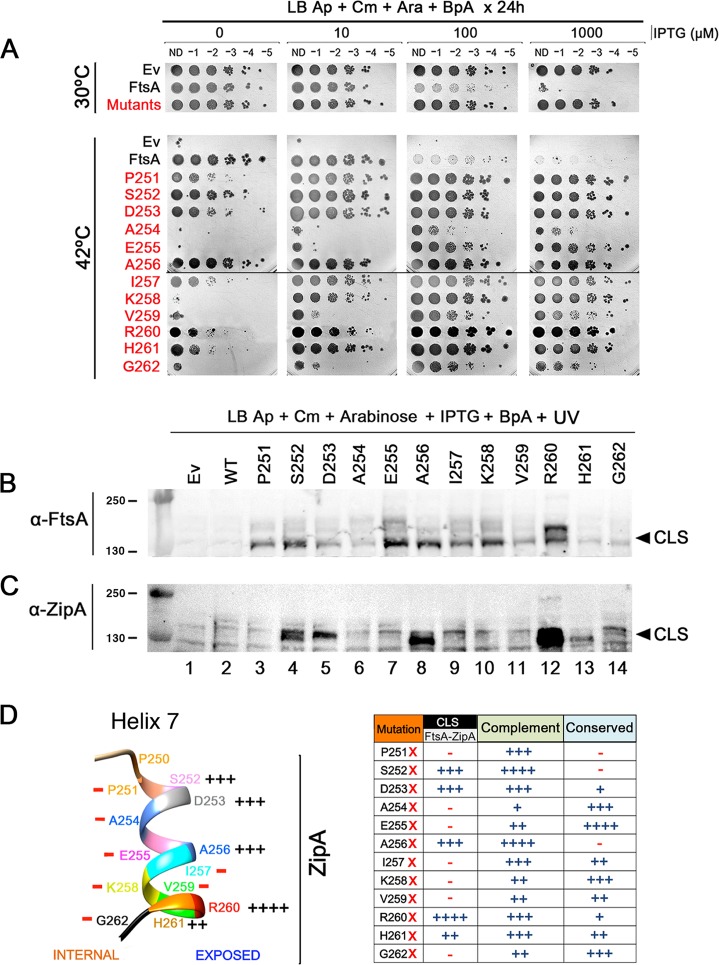
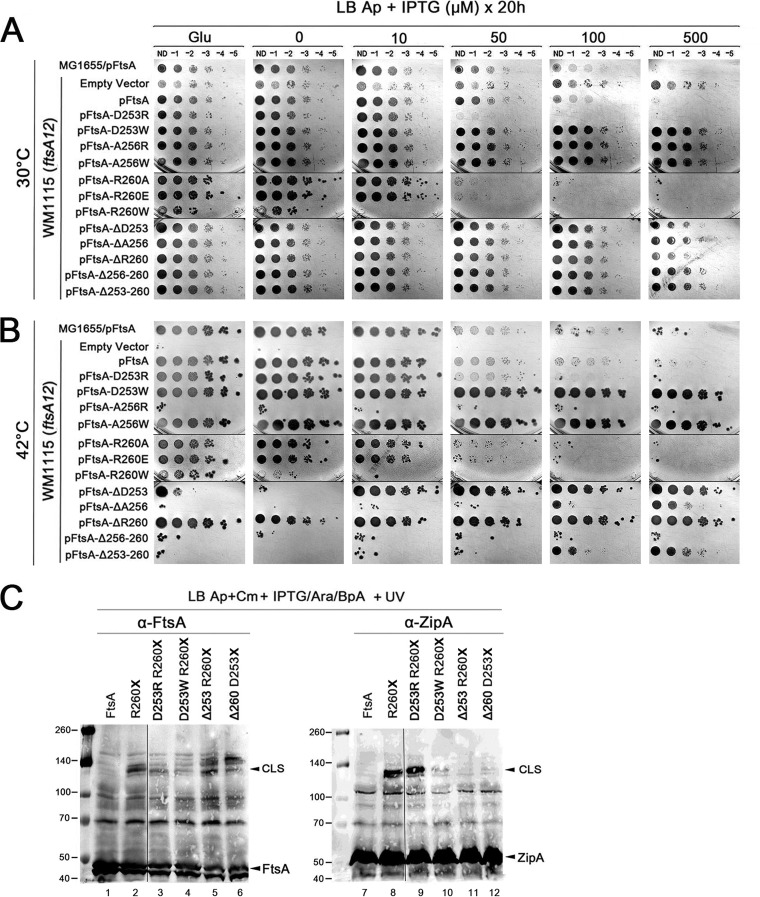
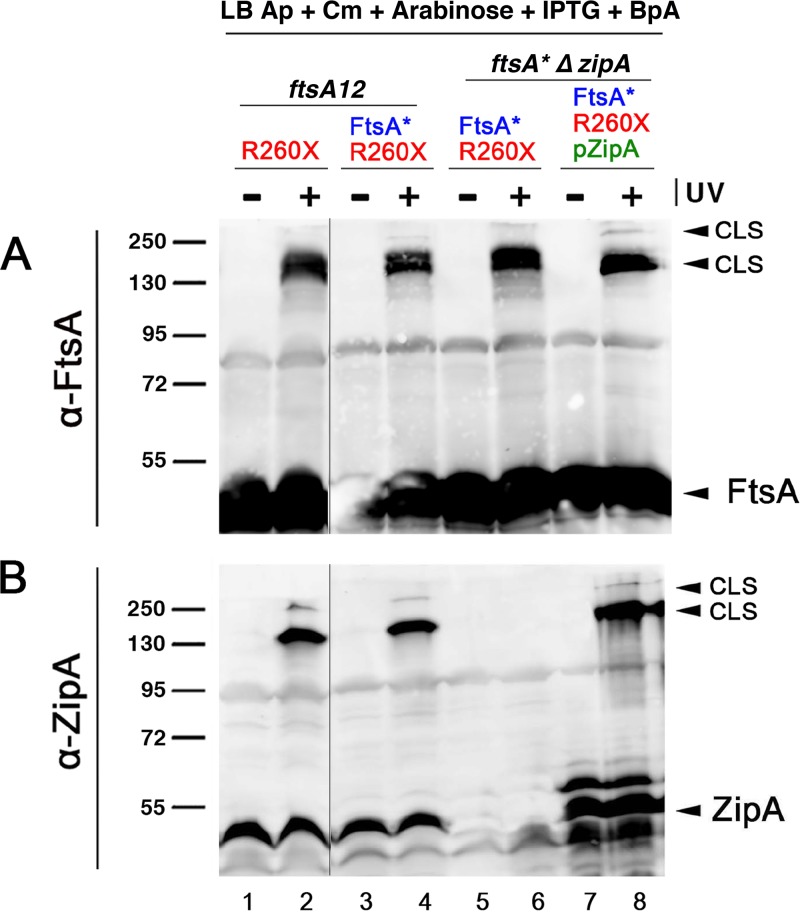
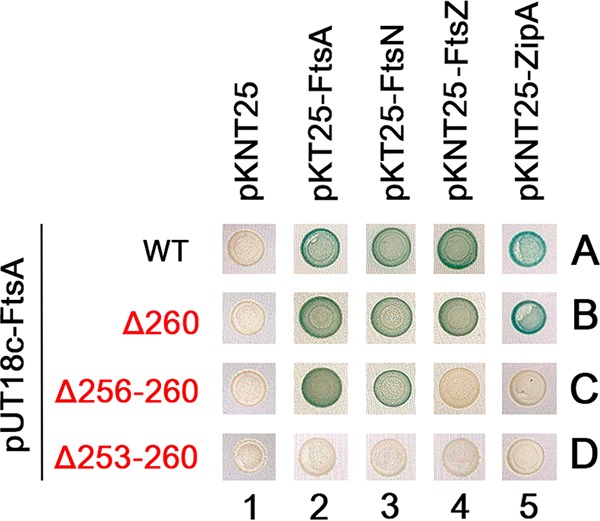
The initiation of Escherichia coli cell division requires three proteins, FtsZ, FtsA, and ZipA, which assemble in a dynamic ring-like structure at midcell. Along with the transmembrane protein ZipA, the actin-like FtsA helps to tether treadmilling polymers of tubulin-like FtsZ to the membrane. In addition to forming homo-oligomers, FtsA and ZipA interact directly with the C-terminal conserved domain of FtsZ. Gain-of-function mutants of FtsA are deficient in forming oligomers and can bypass the need for ZipA, suggesting that ZipA may normally function to disrupt FtsA oligomers, although no direct interaction between FtsA and ZipA has been reported. Here, we use in vivo cross-linking to show that FtsA and ZipA indeed interact directly. We identify the exposed surface of FtsA helix 7, which also participates in binding to ATP through its internal surface, as a key interface needed for the interaction with ZipA. This interaction suggests that FtsZ's membrane tethers may regulate each other's activities.IMPORTANCE To divide, most bacteria first construct a protein machine at the plane of division and then recruit the machinery that will synthesize the division septum. In Escherichia coli, this first stage involves the assembly of FtsZ polymers at midcell, which directly bind to membrane-associated proteins FtsA and ZipA to form a discontinuous ring structure. Although FtsZ directly binds both FtsA and ZipA, it is unclear why FtsZ requires two separate membrane tethers. Here, we uncover a new direct interaction between the tethers, which involves a helix within FtsA that is adjacent to its ATP binding pocket. Our findings imply that in addition to their known roles as FtsZ membrane anchors, FtsA and ZipA may regulate each other's structure and function.
Keywords: Escherichia coli; cell division; cross-linking; ftsA; ftsZ; zipA.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Escherichia coli ZipA Organizes FtsZ Polymers into Dynamic Ring-Like Protofilament Structures.mBio. 2018 Jun 19;9(3):e01008-18. doi: 10.1128/mBio.01008-18. mBio. 2018. PMID: 29921670 Free PMC article.
-
Peptide Linkers within the Essential FtsZ Membrane Tethers ZipA and FtsA Are Nonessential for Cell Division.J Bacteriol. 2020 Feb 25;202(6):e00720-19. doi: 10.1128/JB.00720-19. Print 2020 Feb 25. J Bacteriol. 2020. PMID: 31871036 Free PMC article.
-
ZipA Uses a Two-Pronged FtsZ-Binding Mechanism Necessary for Cell Division.mBio. 2021 Dec 21;12(6):e0252921. doi: 10.1128/mbio.02529-21. Epub 2021 Dec 14. mBio. 2021. PMID: 34903053 Free PMC article.
-
Anchors: A way for FtsZ filaments to stay membrane bound.Mol Microbiol. 2023 Oct;120(4):525-538. doi: 10.1111/mmi.15067. Epub 2023 Apr 28. Mol Microbiol. 2023. PMID: 37503768 Free PMC article. Review.
-
E. coli Cell Cycle Machinery.Subcell Biochem. 2017;84:27-65. doi: 10.1007/978-3-319-53047-5_2. Subcell Biochem. 2017. PMID: 28500522 Review.
Cited by
-
Building the Bacterial Divisome at the Septum.Subcell Biochem. 2024;104:49-71. doi: 10.1007/978-3-031-58843-3_4. Subcell Biochem. 2024. PMID: 38963483 Review.
-
Insights into the assembly and regulation of the bacterial divisome.Nat Rev Microbiol. 2024 Jan;22(1):33-45. doi: 10.1038/s41579-023-00942-x. Epub 2023 Jul 31. Nat Rev Microbiol. 2024. PMID: 37524757 Free PMC article. Review.
-
Localization, Assembly, and Activation of the Escherichia coli Cell Division Machinery.EcoSal Plus. 2021 Dec 15;9(2):eESP00222021. doi: 10.1128/ecosalplus.ESP-0022-2021. Epub 2021 Dec 13. EcoSal Plus. 2021. PMID: 34910577 Free PMC article. Review.
-
Role of the antiparallel double-stranded filament form of FtsA in activating the Escherichia coli divisome.bioRxiv [Preprint]. 2024 Jun 30:2024.06.24.600433. doi: 10.1101/2024.06.24.600433. bioRxiv. 2024. Update in: mBio. 2024 Aug 14;15(8):e0168724. doi: 10.1128/mbio.01687-24. PMID: 38979378 Free PMC article. Updated. Preprint.
-
PlrA (MSMEG_5223) is an essential polar growth regulator in Mycobacterium smegmatis.PLoS One. 2023 Jan 12;18(1):e0280336. doi: 10.1371/journal.pone.0280336. eCollection 2023. PLoS One. 2023. PMID: 36634117 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
