

Phosphorylation at the D53 but Not the T65 Residue of CovR Determines the Repression of rgg and speB Transcription in emm 1- and emm 49-Type Group A Streptococci
- PMID: 30478086
- PMCID: PMC6351742
- DOI: 10.1128/JB.00681-18
Phosphorylation at the D53 but Not the T65 Residue of CovR Determines the Repression of rgg and speB Transcription in emm 1- and emm 49-Type Group A Streptococci
Abstract
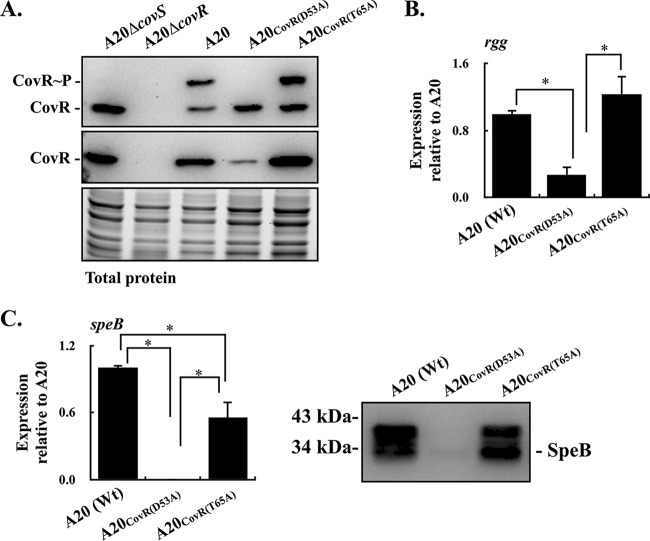
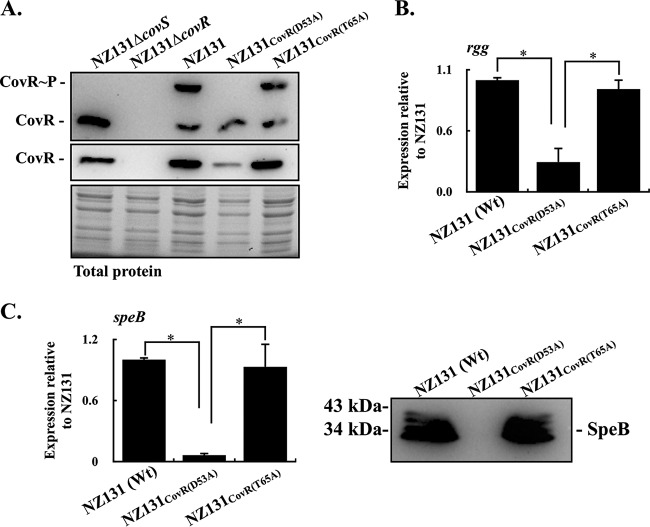
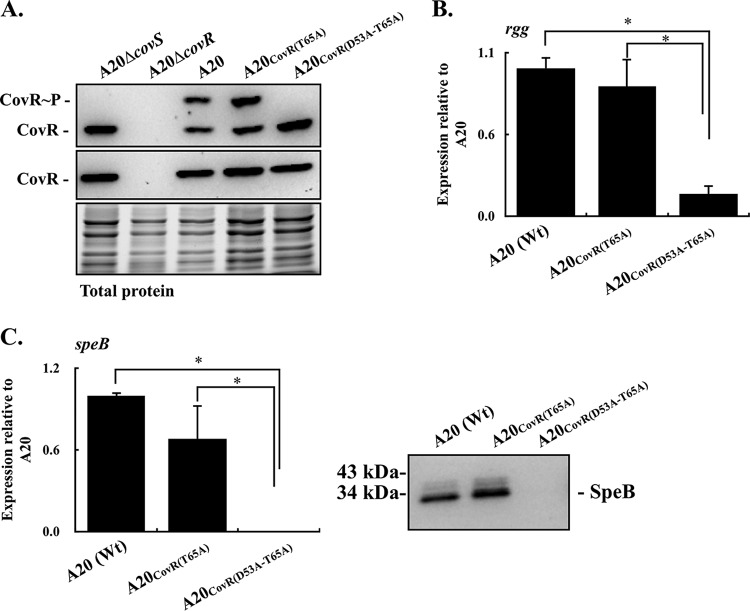
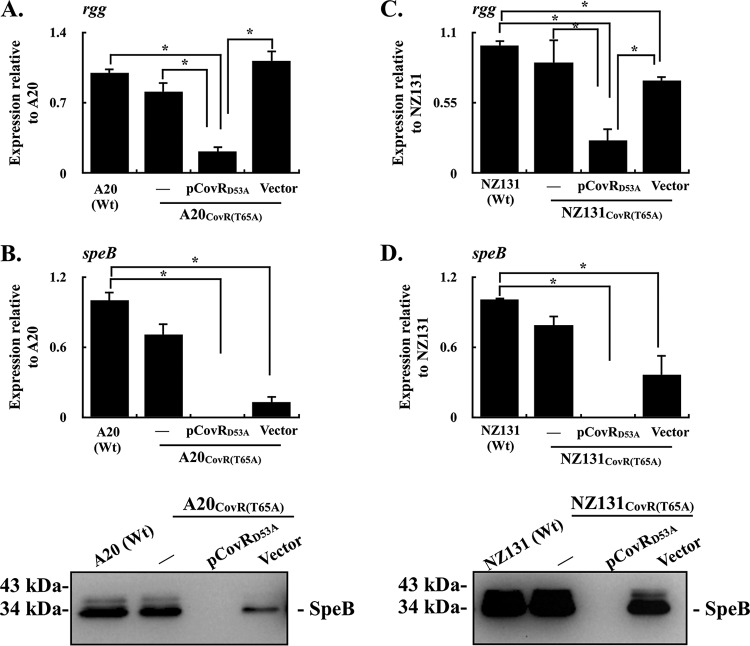
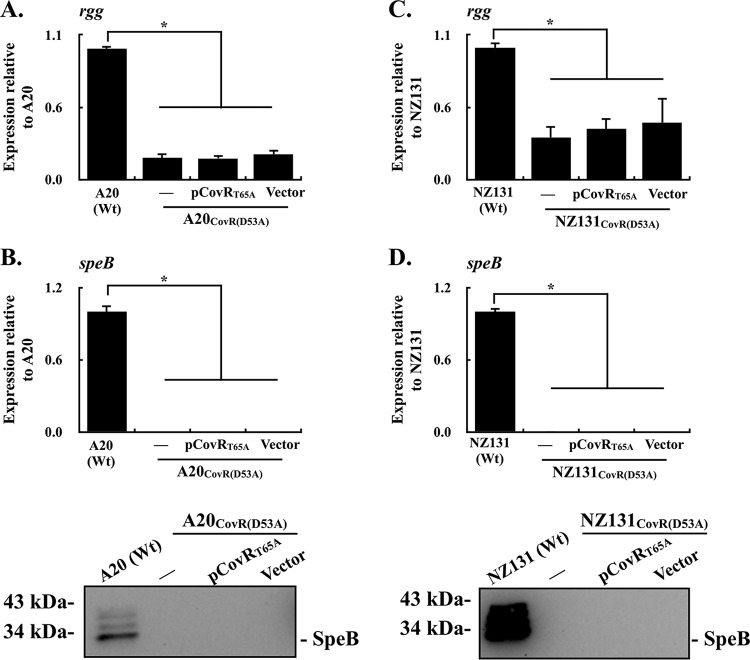
CovR/CovS is a two-component regulatory system in group A Streptococcus and primarily acts as a transcriptional repressor. The D53 residue of CovR (CovRD53) is phosphorylated by the sensor kinase CovS, and the phosphorylated CovRD53 protein binds to the intergenic region of rgg-speB to inhibit speB transcription. Nonetheless, the transcription of rgg and speB is suppressed in covS mutants. The T65 residue of CovR is phosphorylated in a CovS-independent manner, and phosphorylation at the D53 and T65 residues of CovR is mutually exclusive. Therefore, how phosphorylation at the D53 and T65 residues of CovR contributes to the regulation of rgg and speB expression was elucidated. The transcription of rgg and speB was suppressed in the strain that cannot phosphorylate the D53 residue of CovR (CovRD53A mutant) but restored to levels similar to those of the wild-type strain in the CovRT65A mutant. Nonetheless, inactivation of the T65 residue phosphorylation in the CovRD53A mutant cannot derepress the rgg and speB transcription, indicating that phosphorylation at the T65 residue of CovR is not required for repressing rgg and speB transcription. Furthermore, trans complementation of the CovRD53A protein in the strain that expresses the phosphorylated CovRD53 resulted in the repression of rgg and speB transcription. Unlike the direct binding of the phosphorylated CovRD53 protein and its inhibition of speB transcription demonstrated previously, the present study showed that inactivation of phosphorylation at the D53 residue of CovR contributes dominantly in suppressing rgg and speB transcription.IMPORTANCE CovR/CovS is a two-component regulatory system in group A Streptococcus (GAS). The D53 residue of CovR is phosphorylated by CovS, and the phosphorylated CovRD53 binds to the rgg-speB intergenic region and acts as the transcriptional repressor. Nonetheless, the transcription of rgg and Rgg-controlled speB is upregulated in the covR mutant but inhibited in the covS mutant. The present study showed that nonphosphorylated CovRD53 protein inhibits rgg and speB transcription in the presence of the phosphorylated CovRD53in vivo, indicating that nonphosphorylated CovRD53 has a dominant role in suppressing rgg transcription. These results reveal the roles of nonphosphorylated CovRD53 in regulating rgg transcription, which could contribute significantly to invasive phenotypes of covS mutants.
Keywords: CovR/CovS; Rgg; SpeB; group A Streptococcus.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Repression of Rgg But Not Upregulation of LacD.1 in emm1-type covS Mutant Mediates the SpeB Repression in Group A Streptococcus.Front Microbiol. 2016 Nov 29;7:1935. doi: 10.3389/fmicb.2016.01935. eCollection 2016. Front Microbiol. 2016. PMID: 27965655 Free PMC article.
-
RocA Regulates Phosphatase Activity of Virulence Sensor CovS of Group A Streptococcus in Growth Phase- and pH-Dependent Manners.mSphere. 2020 May 20;5(3):e00361-20. doi: 10.1128/mSphere.00361-20. mSphere. 2020. PMID: 32434842 Free PMC article.
-
Dual-site phosphorylation of the control of virulence regulator impacts group a streptococcal global gene expression and pathogenesis.PLoS Pathog. 2014 May 1;10(5):e1004088. doi: 10.1371/journal.ppat.1004088. eCollection 2014 May. PLoS Pathog. 2014. PMID: 24788524 Free PMC article.
-
The two faces of Janus: virulence gene regulation by CovR/S in group A streptococci.Mol Microbiol. 2007 Apr;64(1):34-41. doi: 10.1111/j.1365-2958.2007.05649.x. Mol Microbiol. 2007. PMID: 17376070 Review.
-
From transcription to activation: how group A streptococcus, the flesh-eating pathogen, regulates SpeB cysteine protease production.Mol Microbiol. 2011 Aug;81(3):588-601. doi: 10.1111/j.1365-2958.2011.07709.x. Epub 2011 Jun 24. Mol Microbiol. 2011. PMID: 21707787 Review.
Cited by
-
A Novel CovS Variant Harbored by a Colonization Strain Reduces Streptococcus pyogenes Virulence.J Bacteriol. 2023 Apr 25;205(4):e0003923. doi: 10.1128/jb.00039-23. Epub 2023 Mar 15. J Bacteriol. 2023. PMID: 36920220 Free PMC article.
-
The group A Streptococcus accessory protein RocA: regulatory activity, interacting partners and influence on disease potential.Mol Microbiol. 2020 Jan;113(1):190-207. doi: 10.1111/mmi.14410. Epub 2019 Nov 11. Mol Microbiol. 2020. PMID: 31660653 Free PMC article.
-
Incidence and Effects of Acquisition of the Phage-Encoded ssa Superantigen Gene in Invasive Group A Streptococcus.Front Microbiol. 2021 Jun 4;12:685343. doi: 10.3389/fmicb.2021.685343. eCollection 2021. Front Microbiol. 2021. PMID: 34149675 Free PMC article.
-
Structural basis underlying the synergism of NADase and SLO during group A Streptococcus infection.Commun Biol. 2023 Jan 31;6(1):124. doi: 10.1038/s42003-023-04502-0. Commun Biol. 2023. PMID: 36721030 Free PMC article.
-
Identification of distinct impacts of CovS inactivation on the transcriptome of acapsular group A streptococci.mSystems. 2023 Aug 31;8(4):e0022723. doi: 10.1128/msystems.00227-23. Epub 2023 Jun 26. mSystems. 2023. PMID: 37358280 Free PMC article.
References
-
- Seale AC, Davies MR, Anampiu K, Morpeth SC, Nyongesa S, Mwarumba S, Smeesters PR, Efstratiou A, Karugutu R, Mturi N, Williams TN, Scott JA, Kariuki S, Dougan G, Berkley JA. 2016. Invasive group A Streptococcus infection among children, rural Kenya. Emerg Infect Dis 22:224–232. doi:10.3201/eid2202.151358. - DOI - PMC - PubMed
-
- Engelthaler DM, Valentine M, Bowers J, Pistole J, Driebe EM, Terriquez J, Nienstadt L, Carroll M, Schumacher M, Ormsby ME, Brady S, Livar E, Yazzie D, Waddell V, Peoples M, Komatsu K, Keim P. 2016. Hypervirulent emm59 clone in invasive group A Streptococcus outbreak, southwestern United States. Emerg Infect Dis 22:734–738. doi:10.3201/eid2204.151582. - DOI - PMC - PubMed
-
- Athey TB, Teatero S, Sieswerda LE, Gubbay JB, Marchand-Austin A, Li A, Wasserscheid J, Dewar K, McGeer A, Williams D, Fittipaldi N. 2016. High incidence of invasive group A Streptococcus disease caused by strains of uncommon emm types in Thunder Bay, Ontario, Canada. J Clin Microbiol 54:83–92. doi:10.1128/JCM.02201-15. - DOI - PMC - PubMed
-
- Tse H, Bao JY, Davies MR, Maamary P, Tsoi HW, Tong AH, Ho TC, Lin CH, Gillen CM, Barnett TC, Chen JH, Lee M, Yam WC, Wong CK, Ong CL, Chan YW, Wu CW, Ng T, Lim WW, Tsang TH, Tse CW, Dougan G, Walker MJ, Lok S, Yuen KY. 2012. Molecular characterization of the 2011 Hong Kong scarlet fever outbreak. J Infect Dis 206:341–351. doi:10.1093/infdis/jis362. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
