

CD19-positive antibody-secreting cells provide immune memory
- PMID: 30478153
- PMCID: PMC6258909
- DOI: 10.1182/bloodadvances.2017015172
CD19-positive antibody-secreting cells provide immune memory
Abstract
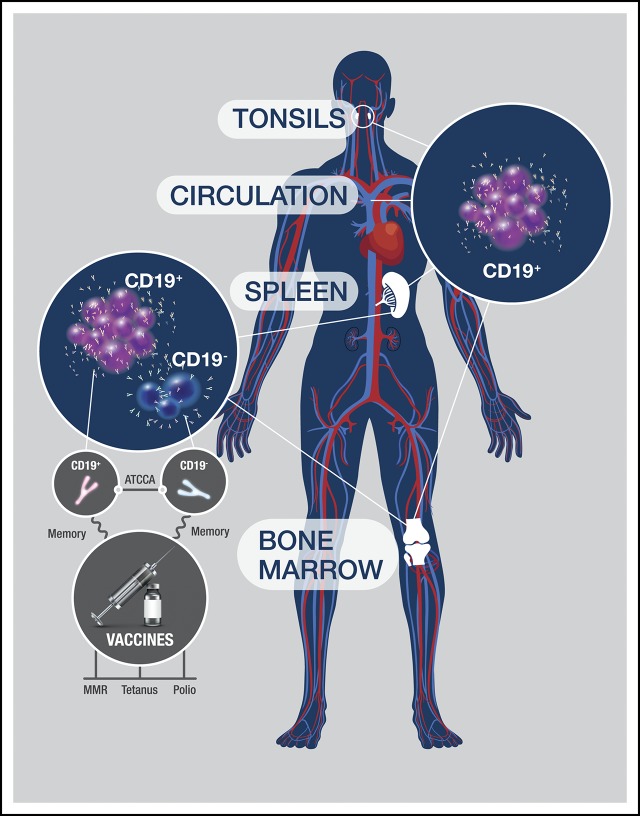
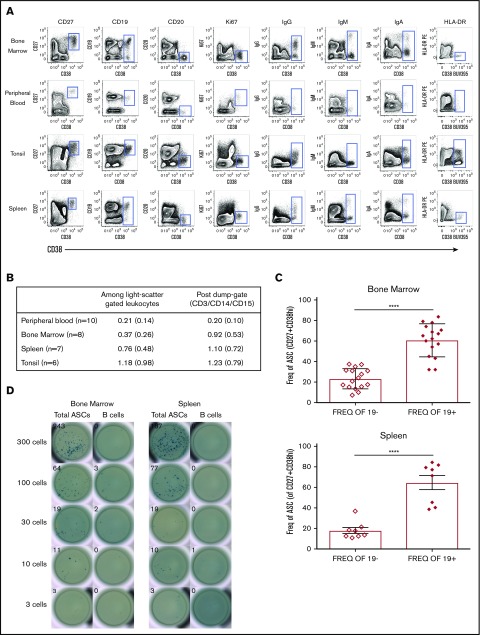
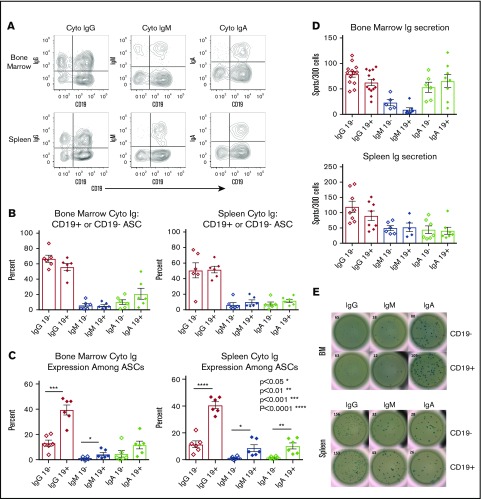
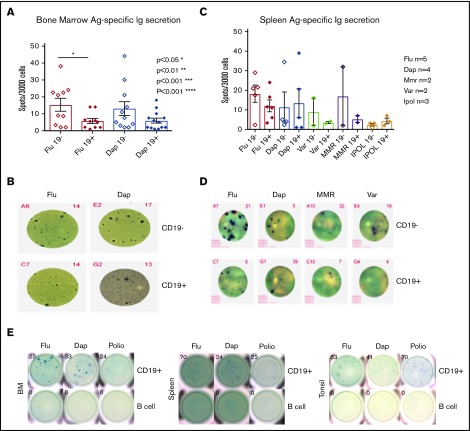
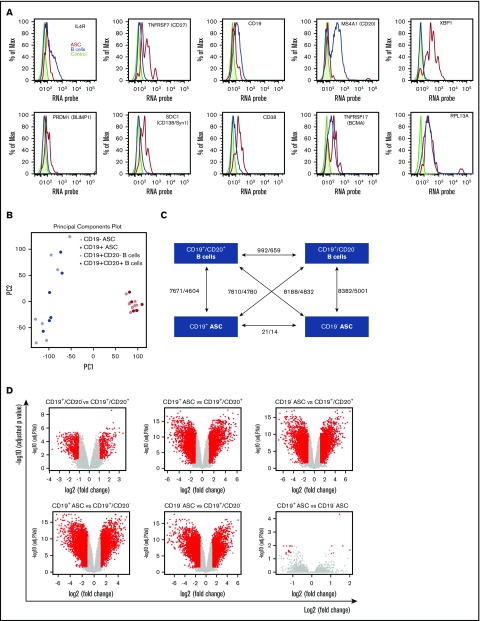
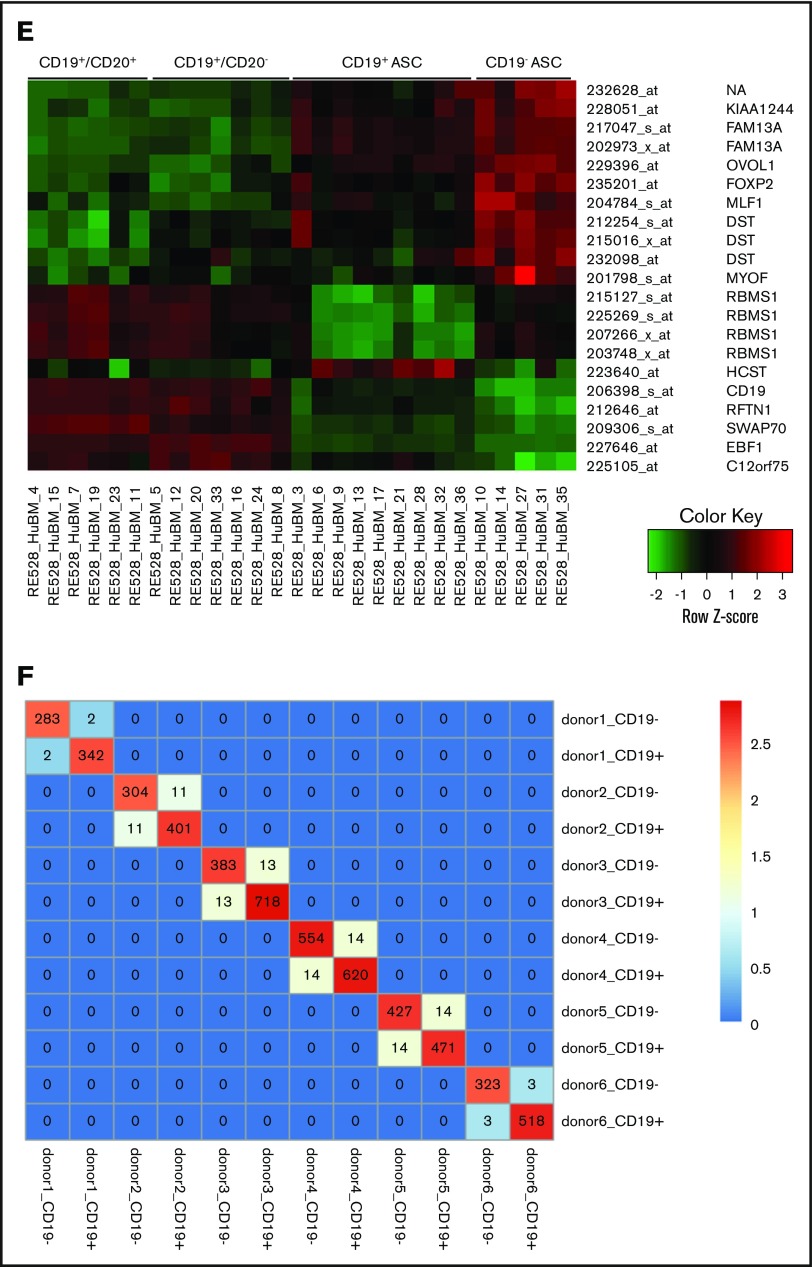
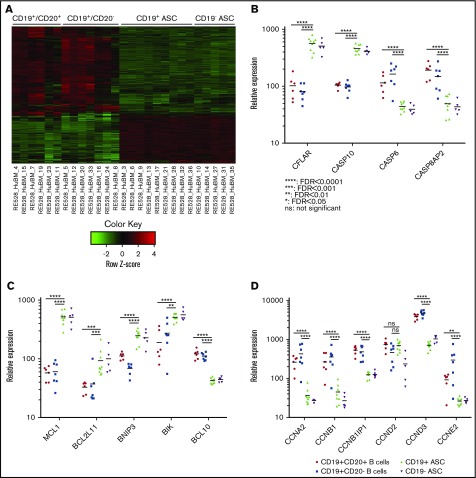
Long-lived antibody-secreting cells (ASCs) are critical for the maintenance of humoral immunity through the continued production of antibodies specific for previously encountered pathogen or vaccine antigens. Recent reports describing humoral immune memory have suggested the importance of long-lived CD19- bone marrow (BM) ASCs, which secrete antibodies recognizing previously encountered vaccine antigens. However, these reports do not agree upon the unique contribution of the CD19+ BM ASC subset toward humoral immunity. Here, we found both CD19+ and negative ASCs from human BM were similar in functional capacity to react to a number of vaccine antigens via ELISpot assays. The CD19+ cells were the predominant ASC population found in lymphoid tissues, and unlike the CD19- ASCs, which were found only in spleen and BM, the CD19+ ASCs were found in tonsil and blood. CD19+ ASCs from the BM, spleen, and tonsil were capable of recognizing polio vaccine antigens, indicating the CD19+ ASC cells play a novel role in long-lasting immune defense. Comparative gene expression analysis indicated CD19+ and negative BM ASCs differed significantly by only 14 distinct messenger RNAs and exhibited similar gene expression for cell cycle, autophagy, and apoptosis control necessary for long life. In addition, we show identical CDR-H3 sequences found on both BM ASC subsets, indicating a shared developmental path. Together, these results provide novel insight for the distribution, function, genetic regulation, and development of long-lived ASCs and may not only impact improved cell therapies but also enhance strategies for vaccine development.
Conflict of interest statement
Conflict-of-interest disclosure: C.J.G., J.C., B.R., C.A.M., R.G., J. Wang, J. Wu, R. Halpin, Y.S., R.R., R.K., Y.W., and R. Herbst are/were full-time employees and shareholders of MedImmune/AstraZeneca.
Figures
References
-
- Ahmed R, Gray D.. Immunological memory and protective immunity: understanding their relation. Science. 1996;272(5258):54-60. - PubMed
-
- Manz RA, Thiel A, Radbruch A. Lifetime of plasma cells in the bone marrow. Nature. 1997;388(6638):133-134. - PubMed
-
- Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunity due to long-lived plasma cells. Immunity. 1998;8(3):363-372. - PubMed
-
- Nutt SL, Hodgkin PD, Tarlinton DM, Corcoran LM. The generation of antibody-secreting plasma cells. Nat Rev Immunol. 2015;15(3):160-171. - PubMed
-
- Shaffer AL, Lin KI, Kuo TC, et al. . Blimp-1 orchestrates plasma cell differentiation by extinguishing the mature B cell gene expression program. Immunity. 2002;17(1):51-62. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
