

Electrophilic fatty acids impair RAD51 function and potentiate the effects of DNA-damaging agents on growth of triple-negative breast cells
- PMID: 30478172
- PMCID: PMC6333886
- DOI: 10.1074/jbc.AC118.005899
Electrophilic fatty acids impair RAD51 function and potentiate the effects of DNA-damaging agents on growth of triple-negative breast cells
Abstract
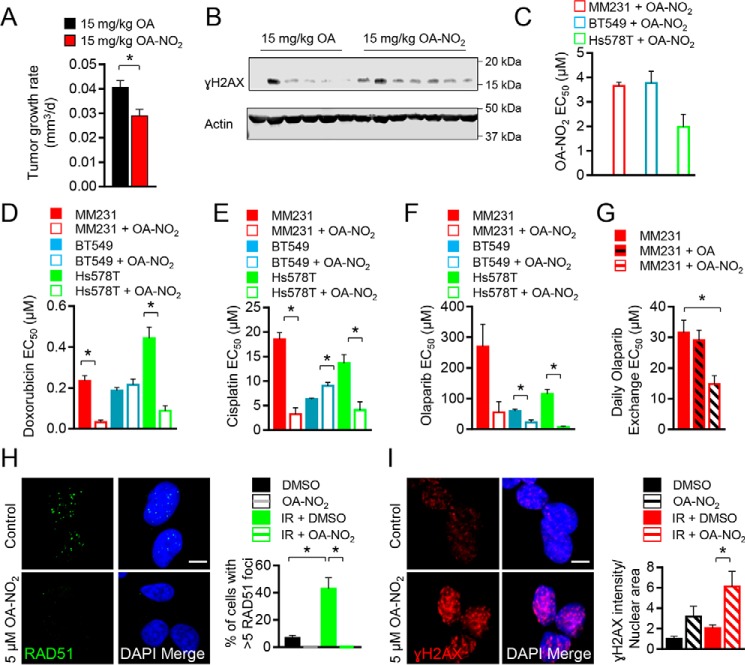
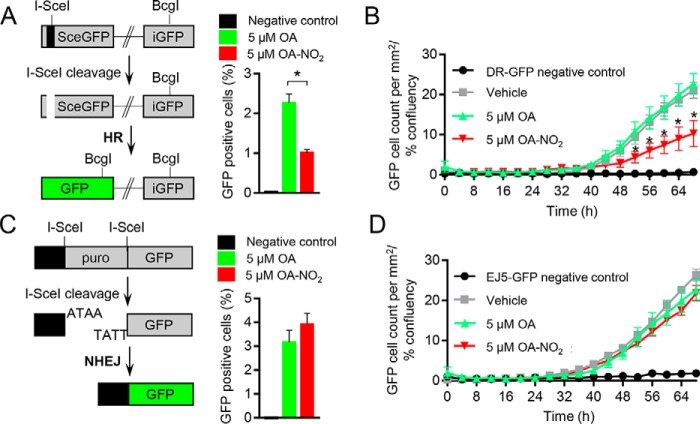
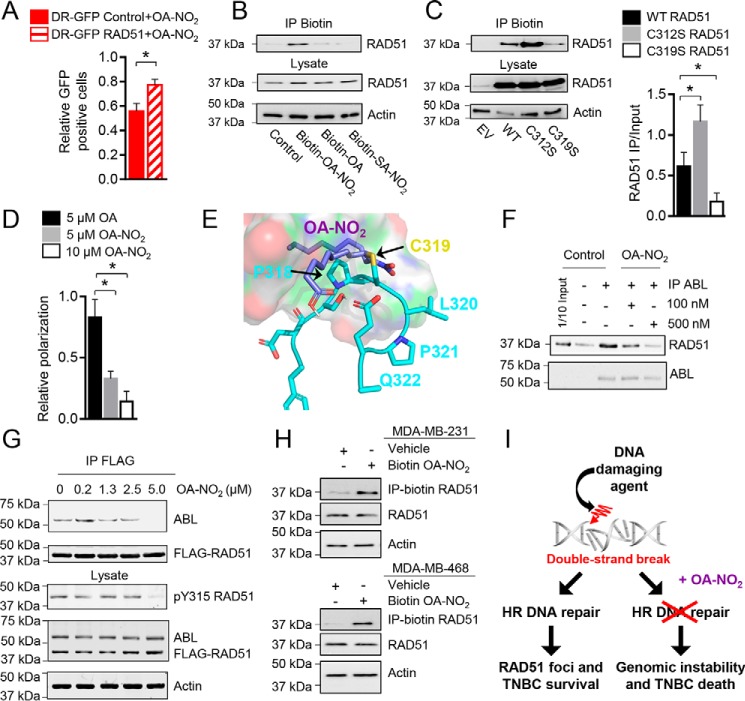
Homologous recombination (HR)-directed DNA double-strand break (DSB) repair enables template-directed DNA repair to maintain genomic stability. RAD51 recombinase (RAD51) is a critical component of HR and facilitates DNA strand exchange in DSB repair. We report here that treating triple-negative breast cancer (TNBC) cells with the fatty acid nitroalkene 10-nitro-octadec-9-enoic acid (OA-NO2) in combination with the antineoplastic DNA-damaging agents doxorubicin, cisplatin, olaparib, and γ-irradiation (IR) enhances the antiproliferative effects of these agents. OA-NO2 inhibited IR-induced RAD51 foci formation and enhanced H2A histone family member X (H2AX) phosphorylation in TNBC cells. Analyses of fluorescent DSB reporter activity with both static-flow cytometry and kinetic live-cell studies enabling temporal resolution of recombination revealed that OA-NO2 inhibits HR and not nonhomologous end joining (NHEJ). OA-NO2 alkylated Cys-319 in RAD51, and this alkylation depended on the Michael acceptor properties of OA-NO2 because nonnitrated and saturated nonelectrophilic analogs of OA-NO2, octadecanoic acid and 10-nitro-octadecanoic acid, did not react with Cys-319. Of note, OA-NO2 alkylation of RAD51 inhibited its binding to ssDNA. RAD51 Cys-319 resides within the SH3-binding site of ABL proto-oncogene 1, nonreceptor tyrosine kinase (ABL1), so we investigated the effect of OA-NO2-mediated Cys-319 alkylation on ABL1 binding and found that OA-NO2 inhibits RAD51-ABL1 complex formation both in vitro and in cell-based immunoprecipitation assays. The inhibition of the RAD51-ABL1 complex also suppressed downstream RAD51 Tyr-315 phosphorylation. In conclusion, RAD51 Cys-319 is a functionally significant site for adduction of soft electrophiles such as OA-NO2 and suggests further investigation of lipid electrophile-based combinational therapies for TNBC.
Keywords: DNA repair; PARP1; Rad51; breast cancer; cysteine-mediated cross-linking; drug action; drug discovery; homologous recombination; nitro fatty acid; nitroalkene; signal transduction; triple negative breast cancer.
© 2019 Asan et al.
Conflict of interest statement
B. A. F. and C.-S. C. W. acknowledge an interest in Complexa, Inc
Figures
References
-
- Woodcock C. C., Huang Y., Woodcock S. R., Salvatore S. R., Singh B., Golin-Bisello F., Davidson N. E., Neumann C. A., Freeman B. A., and Wendell S. G. (2018) Nitro-fatty acid inhibition of triple-negative breast cancer cell viability, migration, invasion, and tumor growth. J. Biol. Chem. 293, 1120–1137 10.1074/jbc.M117.814368 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
