

Iron, erythropoietin, and inflammation regulate hepcidin in Bmp2-deficient mice, but serum iron fails to induce hepcidin in Bmp6-deficient mice
- PMID: 30478858
- PMCID: PMC6492930
- DOI: 10.1002/ajh.25366
Iron, erythropoietin, and inflammation regulate hepcidin in Bmp2-deficient mice, but serum iron fails to induce hepcidin in Bmp6-deficient mice
Abstract
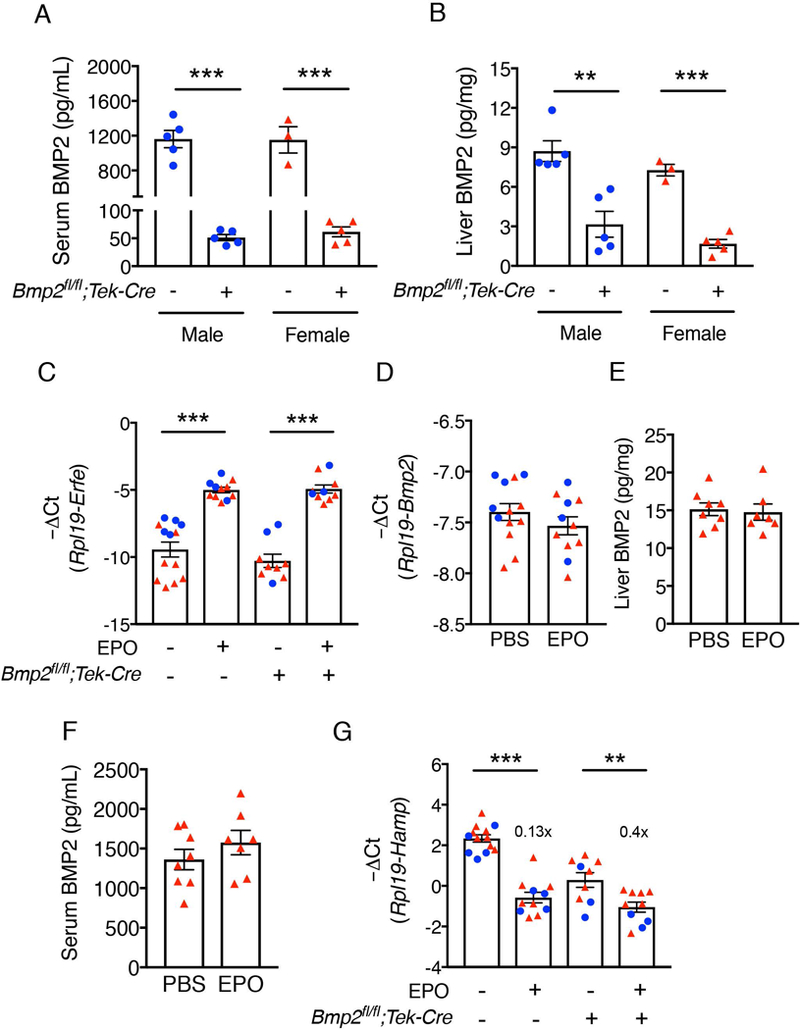
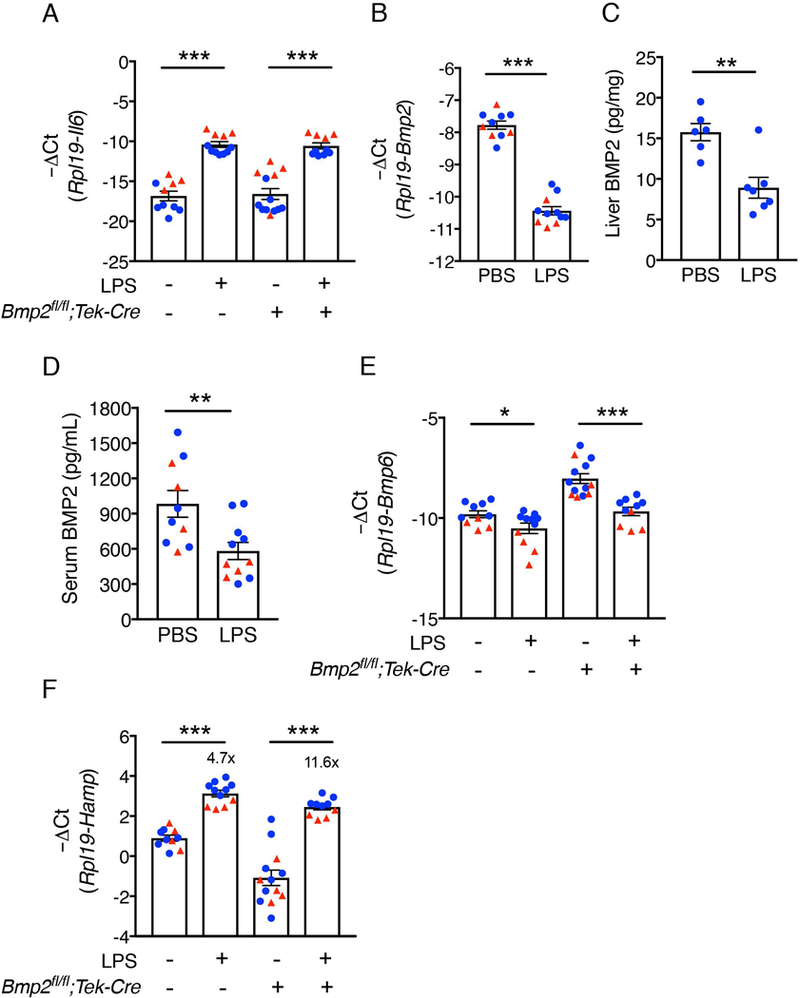
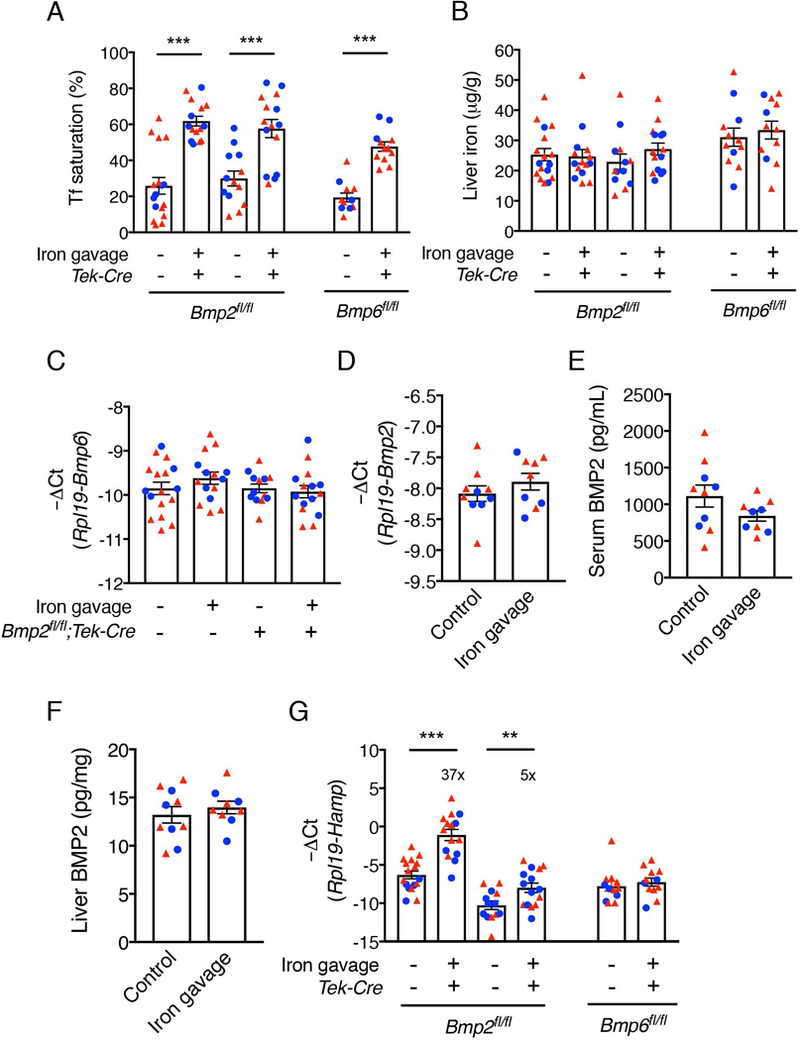
The bone morphogenetic protein (BMP)-SMAD signaling pathway is a key transcriptional regulator of hepcidin in response to tissue iron stores, serum iron, erythropoietic drive and inflammation to increase the iron supply when needed for erythropoiesis, but to prevent the toxicity of iron excess. Recently, BMP2 was reported to play a non-redundant role in hepcidin regulation in addition to BMP6. Here, we used a newly validated BMP2 ELISA assay and mice with a global or endothelial conditional knockout (CKO) of Bmp2 or Bmp6 to examine how BMP2 is regulated and functionally contributes to hepcidin regulation by its major stimuli. Erythropoietin (EPO) did not influence BMP2 expression in control mice, and still suppressed hepcidin in Bmp2 CKO mice. Lipopolysaccharide (LPS) reduced BMP2 expression in control mice, but still induced hepcidin in Bmp2 CKO mice. Chronic dietary iron loading that increased liver iron induced BMP2 expression, whereas acute oral iron gavage that increased serum iron without influencing liver iron did not impact BMP2. However, hepcidin was still induced by both iron loading methods in Bmp2 CKO mice, although the degree of hepcidin induction was blunted relative to control mice. Conversely, acute oral iron gavage failed to induce hepcidin in Bmp6 -/- or CKO mice. Thus, BMP2 has at least a partially redundant role in hepcidin regulation by serum iron, tissue iron, inflammation and erythropoietic drive. In contrast, BMP6 is absolutely required for hepcidin regulation by serum iron.
© 2018 Wiley Periodicals, Inc.
Conflict of interest statement
CONFLICT OF INTEREST DISCLOSURES
JLB has ownership interest in Ferrumax Pharmaceuticals, and has received consulting fees from Keryx Biopharmaceuticals and Disc Medicine. All other authors have nothing to declare.
Figures

Similar articles
-
Endothelial Bone Morphogenetic Protein 2 (Bmp2) Knockout Exacerbates Hemochromatosis in Homeostatic Iron Regulator (Hfe) Knockout Mice but not Bmp6 Knockout Mice.Hepatology. 2020 Aug;72(2):642-655. doi: 10.1002/hep.31048. Epub 2020 May 22. Hepatology. 2020. PMID: 31778583 Free PMC article.
-
BMP5 contributes to hepcidin regulation and systemic iron homeostasis in mice.Blood. 2023 Oct 12;142(15):1312-1322. doi: 10.1182/blood.2022019195. Blood. 2023. PMID: 37478395 Free PMC article.
-
Bone morphogenetic protein 2 controls iron homeostasis in mice independent of Bmp6.Am J Hematol. 2017 Nov;92(11):1204-1213. doi: 10.1002/ajh.24888. Epub 2017 Sep 25. Am J Hematol. 2017. PMID: 28815688 Free PMC article.
-
Hepcidin and the BMP-SMAD pathway: An unexpected liaison.Vitam Horm. 2019;110:71-99. doi: 10.1016/bs.vh.2019.01.004. Epub 2019 Feb 10. Vitam Horm. 2019. PMID: 30798817 Review.
-
Bone morphogenic proteins in iron homeostasis.Bone. 2020 Sep;138:115495. doi: 10.1016/j.bone.2020.115495. Epub 2020 Jun 23. Bone. 2020. PMID: 32585319 Free PMC article. Review.
Cited by
-
Coordination of iron homeostasis by bone morphogenetic proteins: Current understanding and unanswered questions.Dev Dyn. 2022 Jan;251(1):26-46. doi: 10.1002/dvdy.372. Epub 2021 May 25. Dev Dyn. 2022. PMID: 33993583 Free PMC article.
-
Endothelial Bone Morphogenetic Protein 2 (Bmp2) Knockout Exacerbates Hemochromatosis in Homeostatic Iron Regulator (Hfe) Knockout Mice but not Bmp6 Knockout Mice.Hepatology. 2020 Aug;72(2):642-655. doi: 10.1002/hep.31048. Epub 2020 May 22. Hepatology. 2020. PMID: 31778583 Free PMC article.
-
Bone morphogenetic protein signaling in inflammation.Exp Biol Med (Maywood). 2019 Feb;244(2):147-156. doi: 10.1177/1535370219828694. Epub 2019 Feb 7. Exp Biol Med (Maywood). 2019. PMID: 30732465 Free PMC article. Review.
-
Effects of Two Kinds of Iron Nanoparticles as Reactive Oxygen Species Inducer and Scavenger on the Transcriptomic Profiles of Two Human Leukemia Cells with Different Stemness.Nanomaterials (Basel). 2020 Sep 30;10(10):1951. doi: 10.3390/nano10101951. Nanomaterials (Basel). 2020. PMID: 33007950 Free PMC article.
-
BMP5 contributes to hepcidin regulation and systemic iron homeostasis in mice.Blood. 2023 Oct 12;142(15):1312-1322. doi: 10.1182/blood.2022019195. Blood. 2023. PMID: 37478395 Free PMC article.
References
-
- Stoltzfus RJ. Iron deficiency: global prevalence and consequences. Food Nutr Bull. 2003;24(4 Suppl):S99–103. - PubMed
-
- McLean E, Cogswell M, Egli I, Wojdyla D, de Benoist B. Worldwide prevalence of anaemia, WHO Vitamin and Mineral Nutrition Information System, 1993–2005. Public Health Nutr. 2009;12(4):444–454. - PubMed
-
- Chevion M A site-specific mechanism for free radical induced biological damage: the essential role of redox-active transition metals. Free Radic Biol Med. 1988;5(1):27–37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials