

Neuronal coding of multiscale temporal features in communication sequences within the bat auditory cortex
- PMID: 30480101
- PMCID: PMC6244232
- DOI: 10.1038/s42003-018-0205-5
Neuronal coding of multiscale temporal features in communication sequences within the bat auditory cortex
Abstract
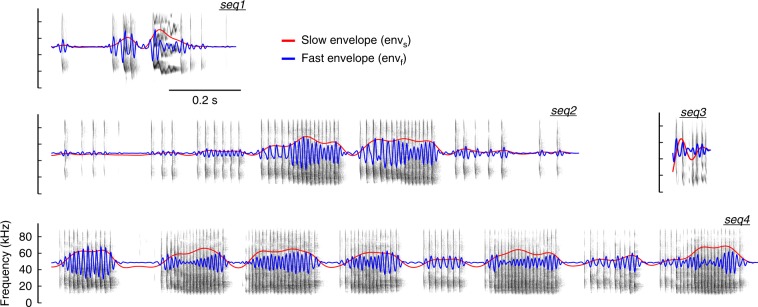
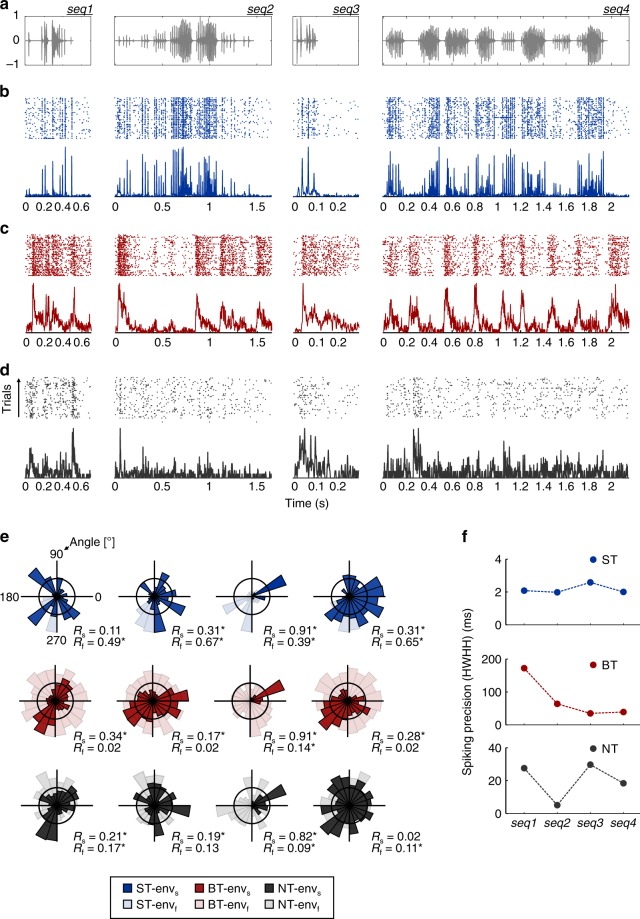
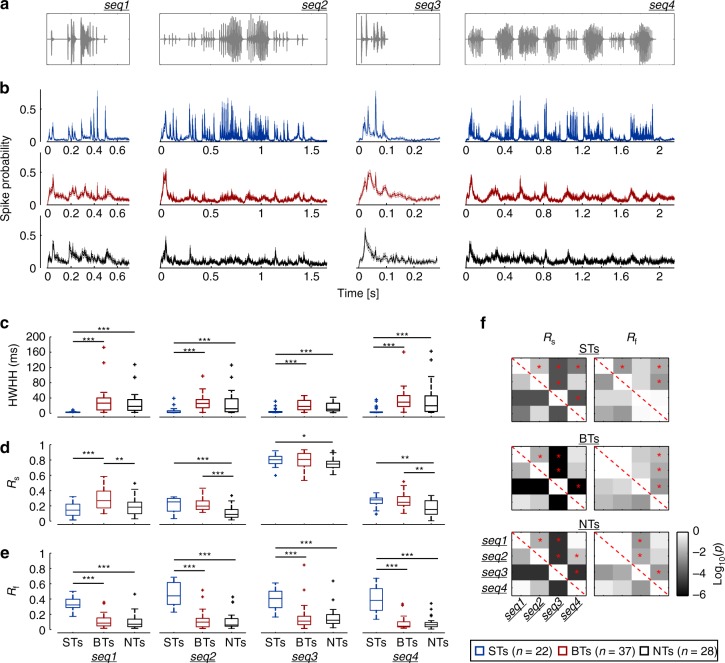
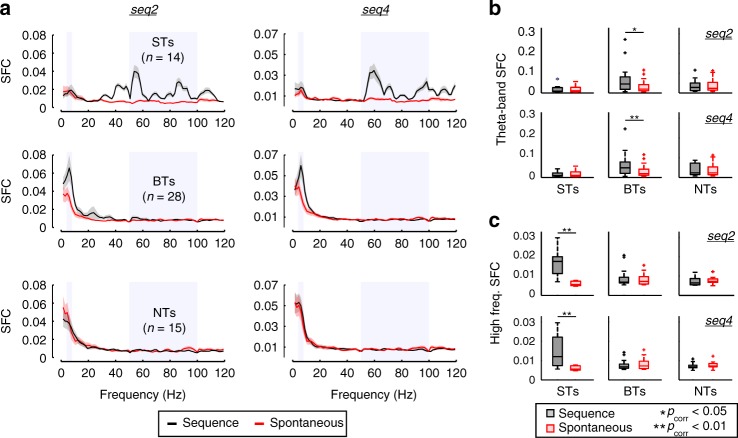
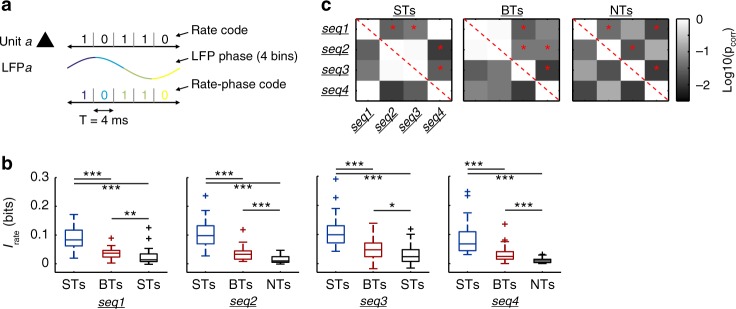
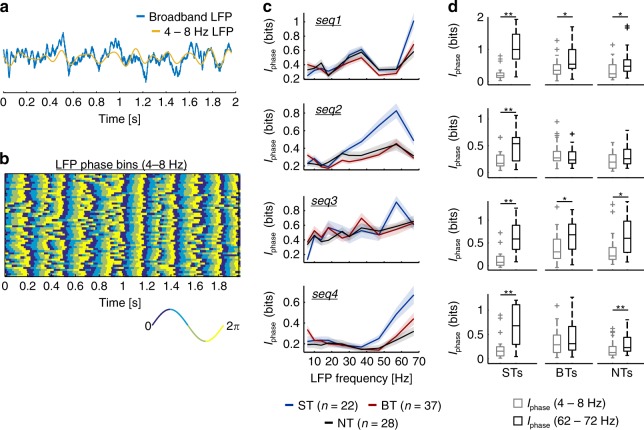
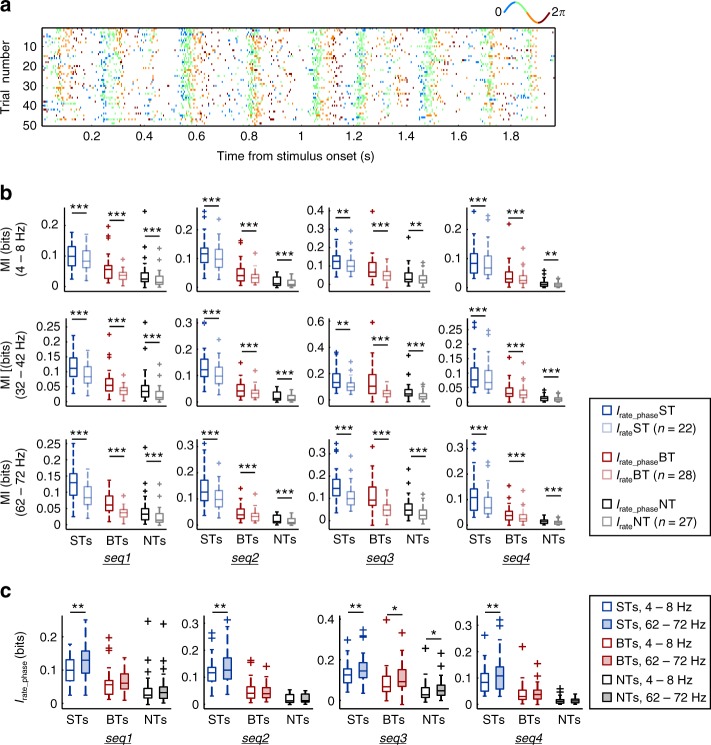
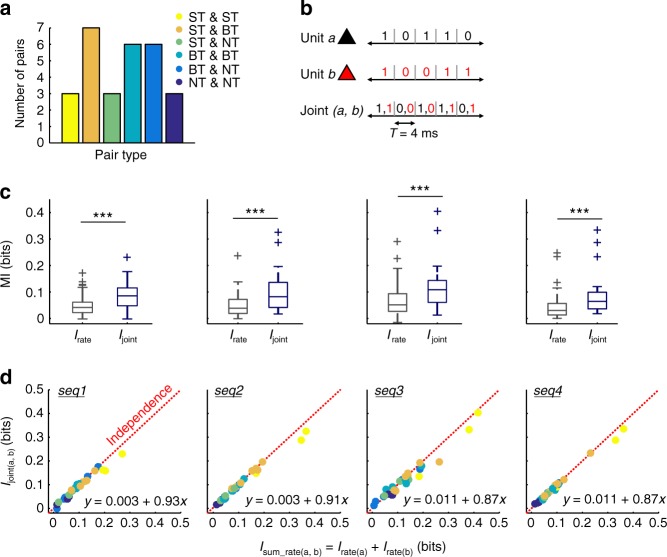
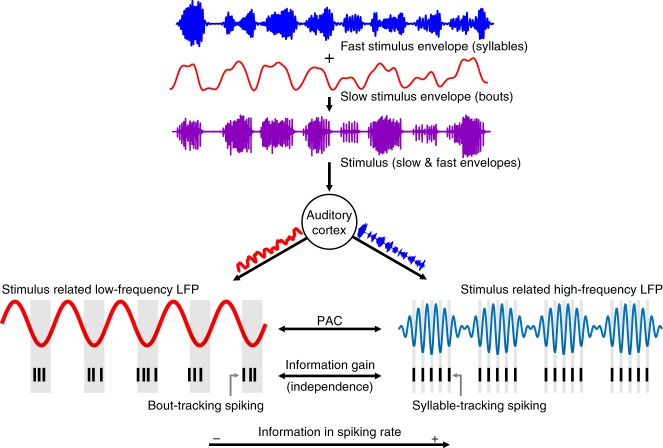
Experimental evidence supports that cortical oscillations represent multiscale temporal modulations existent in natural stimuli, yet little is known about the processing of these multiple timescales at a neuronal level. Here, using extracellular recordings from the auditory cortex (AC) of awake bats (Carollia perspicillata), we show the existence of three neuronal types which represent different levels of the temporal structure of conspecific vocalizations, and therefore constitute direct evidence of multiscale temporal processing of naturalistic stimuli by neurons in the AC. These neuronal subpopulations synchronize differently to local-field potentials, particularly in theta- and high frequency bands, and are informative to a different degree in terms of their spike rate. Interestingly, we also observed that both low and high frequency cortical oscillations can be highly informative about the listened calls. Our results suggest that multiscale neuronal processing allows for the precise and non-redundant representation of natural vocalizations in the AC.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Laminar specificity of oscillatory coherence in the auditory cortex.Brain Struct Funct. 2019 Nov;224(8):2907-2924. doi: 10.1007/s00429-019-01944-3. Epub 2019 Aug 27. Brain Struct Funct. 2019. PMID: 31456067
-
Low-Frequency Spike-Field Coherence Is a Fingerprint of Periodicity Coding in the Auditory Cortex.iScience. 2018 Nov 30;9:47-62. doi: 10.1016/j.isci.2018.10.009. Epub 2018 Oct 16. iScience. 2018. PMID: 30384133 Free PMC article.
-
Processing of fast amplitude modulations in bat auditory cortex matches communication call-specific sound features.J Neurophysiol. 2019 Apr 1;121(4):1501-1512. doi: 10.1152/jn.00748.2018. Epub 2019 Feb 20. J Neurophysiol. 2019. PMID: 30785811
-
How do auditory cortex neurons represent communication sounds?Hear Res. 2013 Nov;305:102-12. doi: 10.1016/j.heares.2013.03.011. Epub 2013 Apr 17. Hear Res. 2013. PMID: 23603138 Review.
-
Coding of communication calls in the subcortical and cortical structures of the auditory system.Physiol Res. 2008;57 Suppl 3:S149-S159. doi: 10.33549/physiolres.931608. Epub 2008 May 13. Physiol Res. 2008. PMID: 18481905 Review.
Cited by
-
The fully automated bat (FAB) flight room: A human-free environment for studying navigation in flying bats and its initial application to the retrosplenial cortex.J Neurosci Methods. 2021 Jan 15;348:108970. doi: 10.1016/j.jneumeth.2020.108970. Epub 2020 Oct 14. J Neurosci Methods. 2021. PMID: 33065152 Free PMC article.
-
Echolocation-related reversal of information flow in a cortical vocalization network.Nat Commun. 2022 Jun 25;13(1):3642. doi: 10.1038/s41467-022-31230-6. Nat Commun. 2022. PMID: 35752629 Free PMC article.
-
Bats distress vocalizations carry fast amplitude modulations that could represent an acoustic correlate of roughness.Sci Rep. 2020 Apr 30;10(1):7332. doi: 10.1038/s41598-020-64323-7. Sci Rep. 2020. PMID: 32355293 Free PMC article.
-
Fronto-Temporal Coupling Dynamics During Spontaneous Activity and Auditory Processing in the Bat Carollia perspicillata.Front Syst Neurosci. 2020 Mar 20;14:14. doi: 10.3389/fnsys.2020.00014. eCollection 2020. Front Syst Neurosci. 2020. PMID: 32265670 Free PMC article.
-
What do mammals have to say about the neurobiology of acoustic communication?Mol Psychol. 2023;2:5. doi: 10.12688/molpsychol.17539.1. Epub 2023 May 4. Mol Psychol. 2023. PMID: 38827277 Free PMC article.
References
LinkOut - more resources
Full Text Sources
