

Enhanced axonal neuregulin-1 type-III signaling ameliorates neurophysiology and hypomyelination in a Charcot-Marie-Tooth type 1B mouse model
- PMID: 30481294
- PMCID: PMC6400047
- DOI: 10.1093/hmg/ddy411
Enhanced axonal neuregulin-1 type-III signaling ameliorates neurophysiology and hypomyelination in a Charcot-Marie-Tooth type 1B mouse model
Abstract
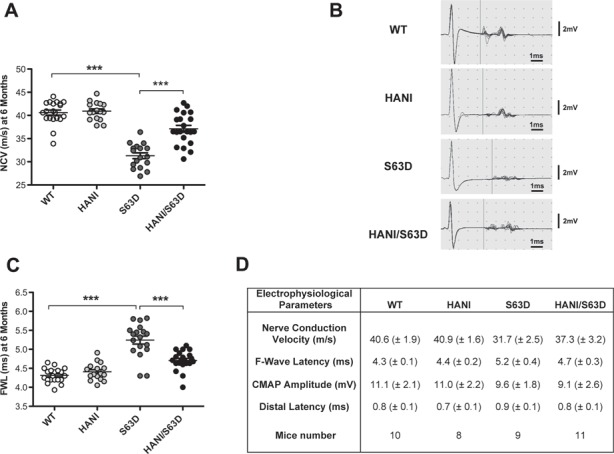
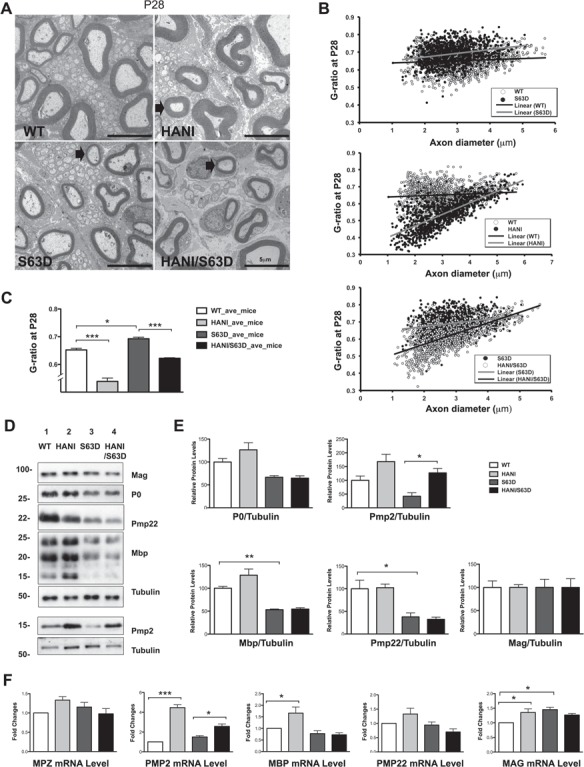
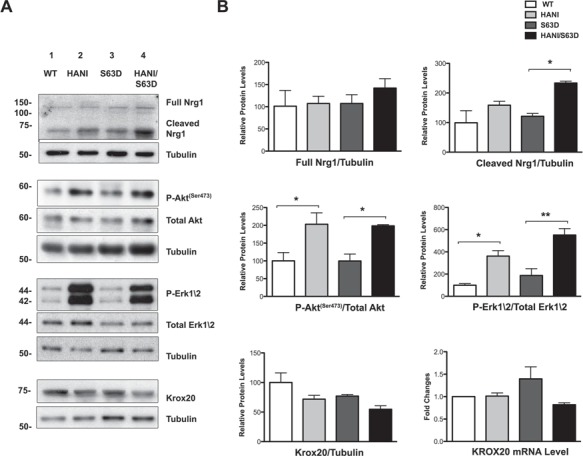
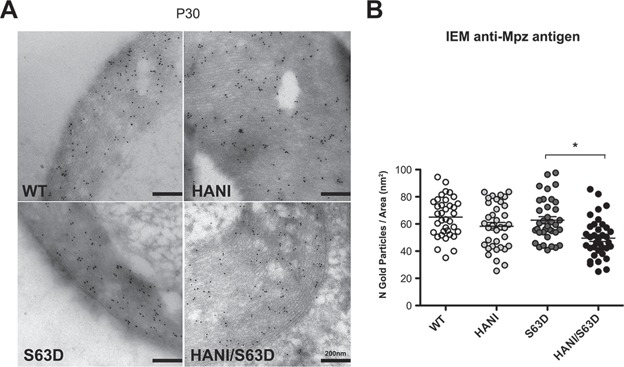
Charcot-Marie-Tooth (CMT) neuropathies are a group of genetic disorders that affect the peripheral nervous system with heterogeneous pathogenesis and no available treatment. Axonal neuregulin 1 type III (Nrg1TIII) drives peripheral nerve myelination by activating downstream signaling pathways such as PI3K/Akt and MAPK/Erk that converge on master transcriptional regulators of myelin genes, such as Krox20. We reasoned that modulating Nrg1TIII activity may constitute a general therapeutic strategy to treat CMTs that are characterized by reduced levels of myelination. Here we show that genetic overexpression of Nrg1TIII ameliorates neurophysiological and morphological parameters in a mouse model of demyelinating CMT1B, without exacerbating the toxic gain-of-function that underlies the neuropathy. Intriguingly, the mechanism appears not to be related to Krox20 or myelin gene upregulation, but rather to a beneficial rebalancing in the stoichiometry of myelin lipids and proteins. Finally, we provide proof of principle that stimulating Nrg1TIII signaling, by pharmacological suppression of the Nrg1TIII inhibitor tumor necrosis factor-alpha-converting enzyme (TACE/ADAM17), also ameliorates the neuropathy. Thus, modulation of Nrg1TIII by TACE/ADAM17 inhibition may represent a general treatment for hypomyelinating neuropathies.
© The Author(s) 2018. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
Similar articles
-
Neuregulin 1 type III improves peripheral nerve myelination in a mouse model of congenital hypomyelinating neuropathy.Hum Mol Genet. 2019 Apr 15;28(8):1260-1273. doi: 10.1093/hmg/ddy420. Hum Mol Genet. 2019. PMID: 30535360 Free PMC article.
-
Activation of XBP1s attenuates disease severity in models of proteotoxic Charcot-Marie-Tooth type 1B.Brain. 2025 Jun 3;148(6):1978-1993. doi: 10.1093/brain/awae407. Brain. 2025. PMID: 39979221 Free PMC article.
-
c-Jun activation in Schwann cells protects against loss of sensory axons in inherited neuropathy.Brain. 2014 Nov;137(Pt 11):2922-37. doi: 10.1093/brain/awu257. Epub 2014 Sep 12. Brain. 2014. PMID: 25216747 Free PMC article.
-
Pathomechanisms of mutant proteins in Charcot-Marie-Tooth disease.Neuromolecular Med. 2006;8(1-2):217-42. doi: 10.1385/nmm:8:1-2:217. Neuromolecular Med. 2006. PMID: 16775378 Review.
-
New insights into signaling during myelination in zebrafish.Curr Top Dev Biol. 2011;97:1-19. doi: 10.1016/B978-0-12-385975-4.00007-3. Curr Top Dev Biol. 2011. PMID: 22074600 Free PMC article. Review.
Cited by
-
The Complex Work of Proteases and Secretases in Wallerian Degeneration: Beyond Neuregulin-1.Front Cell Neurosci. 2019 Mar 20;13:93. doi: 10.3389/fncel.2019.00093. eCollection 2019. Front Cell Neurosci. 2019. PMID: 30949030 Free PMC article. Review.
-
Animal Models as a Tool to Design Therapeutical Strategies for CMT-like Hereditary Neuropathies.Brain Sci. 2021 Sep 18;11(9):1237. doi: 10.3390/brainsci11091237. Brain Sci. 2021. PMID: 34573256 Free PMC article. Review.
-
Imbalance of NRG1-ERBB2/3 signalling underlies altered myelination in Charcot-Marie-Tooth disease 4H.Brain. 2023 May 2;146(5):1844-1858. doi: 10.1093/brain/awac402. Brain. 2023. PMID: 36314052 Free PMC article.
-
Navigating the Landscape of CMT1B: Understanding Genetic Pathways, Disease Models, and Potential Therapeutic Approaches.Int J Mol Sci. 2024 Aug 26;25(17):9227. doi: 10.3390/ijms25179227. Int J Mol Sci. 2024. PMID: 39273178 Free PMC article. Review.
-
Pmp2+ Schwann Cells Maintain the Survival of Large-Caliber Motor Axons.J Neurosci. 2025 Mar 26;45(13):e1362242025. doi: 10.1523/JNEUROSCI.1362-24.2025. J Neurosci. 2025. PMID: 39880678
References
-
- Jessen K.R. and Mirsky R. (2005) The origin and development of glial cells in peripheral nerves. Nat. Rev. Neurosci., 6, 671–682. - PubMed
-
- Nave K.A. (2010) Myelination and support of axonal integrity by glia. Nature, 468, 244–252. - PubMed
-
- Barisic N., Claeys K.G., Sirotkovic-Skerlev M., Lofgren A., Nelis E., De Jonghe P. and Timmerman V. (2008) Charcot–Marie–Tooth disease: a clinico-genetic confrontation. Ann. Hum. Genet., 72, 416–441. - PubMed
-
- Reilly M.M. and Shy M.E. (2009) Diagnosis and new treatments in genetic neuropathies. J. Neurol. Neurosurg. Psychiatry, 80, 1304–1314. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
