

Nuclear genome sequence of the plastid-lacking cryptomonad Goniomonas avonlea provides insights into the evolution of secondary plastids
- PMID: 30482201
- PMCID: PMC6260743
- DOI: 10.1186/s12915-018-0593-5
Nuclear genome sequence of the plastid-lacking cryptomonad Goniomonas avonlea provides insights into the evolution of secondary plastids
Abstract
Background: The evolution of photosynthesis has been a major driver in eukaryotic diversification. Eukaryotes have acquired plastids (chloroplasts) either directly via the engulfment and integration of a photosynthetic cyanobacterium (primary endosymbiosis) or indirectly by engulfing a photosynthetic eukaryote (secondary or tertiary endosymbiosis). The timing and frequency of secondary endosymbiosis during eukaryotic evolution is currently unclear but may be resolved in part by studying cryptomonads, a group of single-celled eukaryotes comprised of both photosynthetic and non-photosynthetic species. While cryptomonads such as Guillardia theta harbor a red algal-derived plastid of secondary endosymbiotic origin, members of the sister group Goniomonadea lack plastids. Here, we present the genome of Goniomonas avonlea-the first for any goniomonad-to address whether Goniomonadea are ancestrally non-photosynthetic or whether they lost a plastid secondarily.
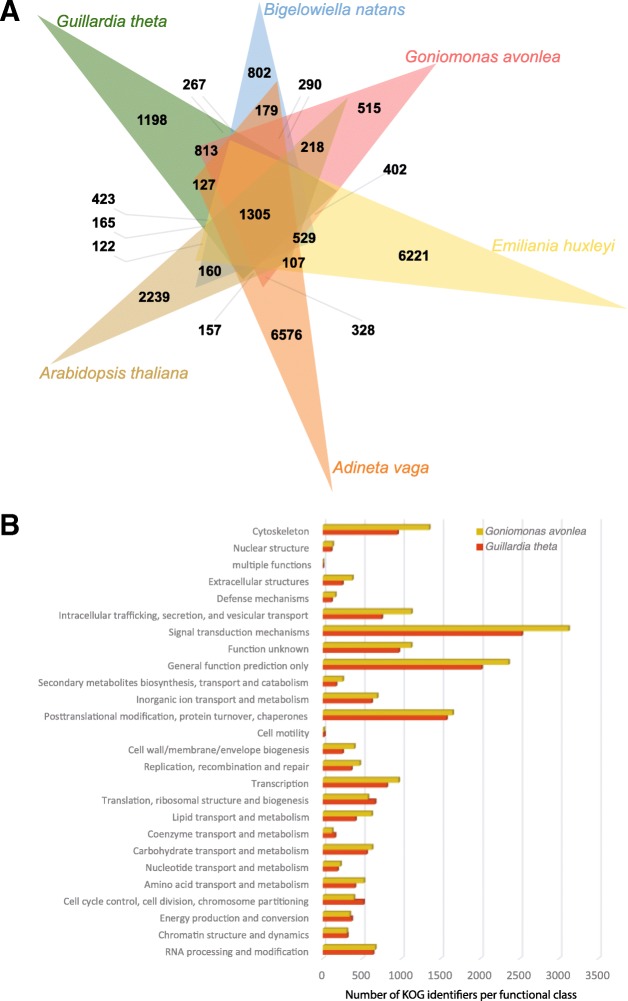
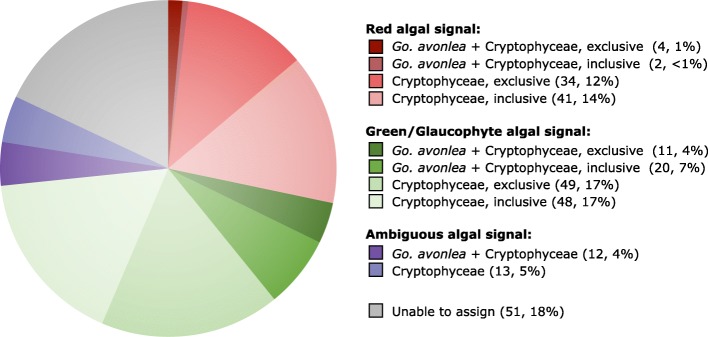
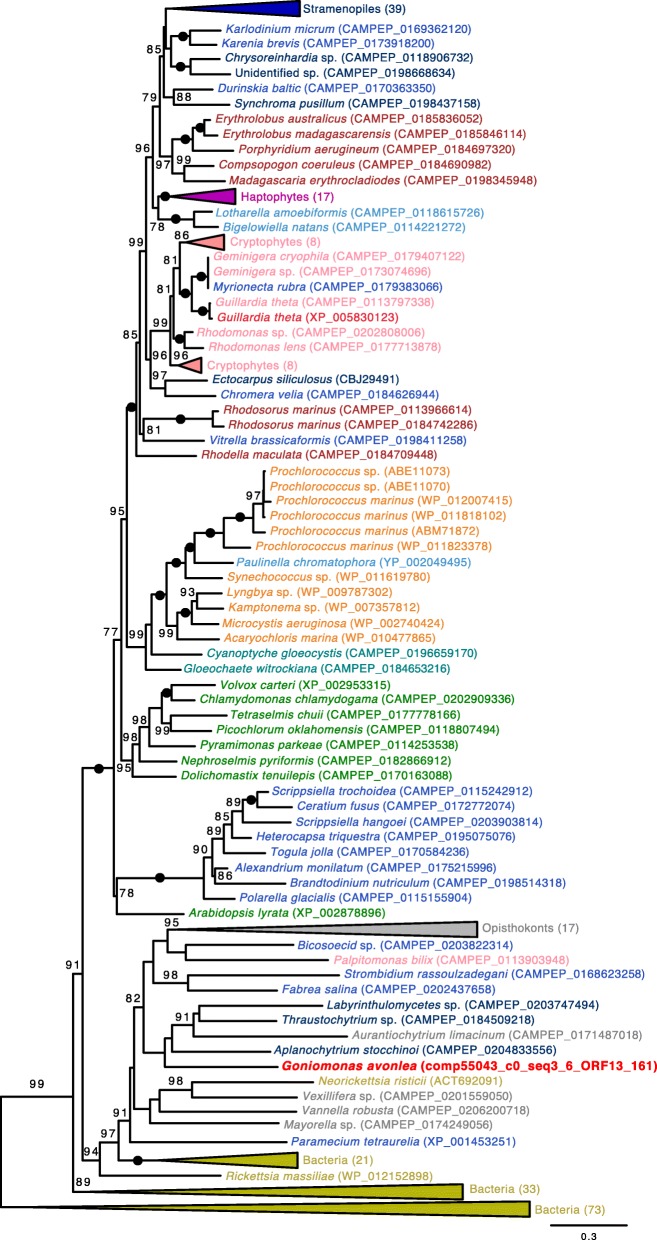
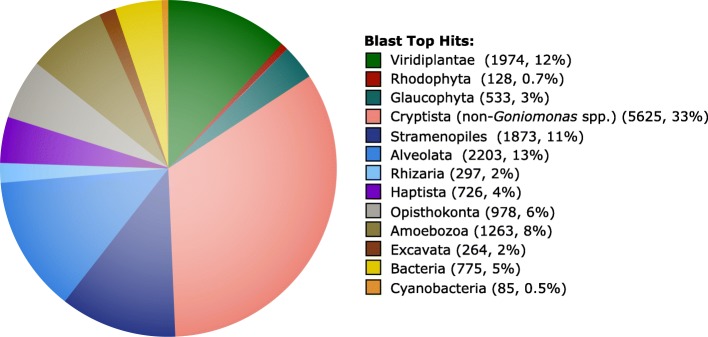
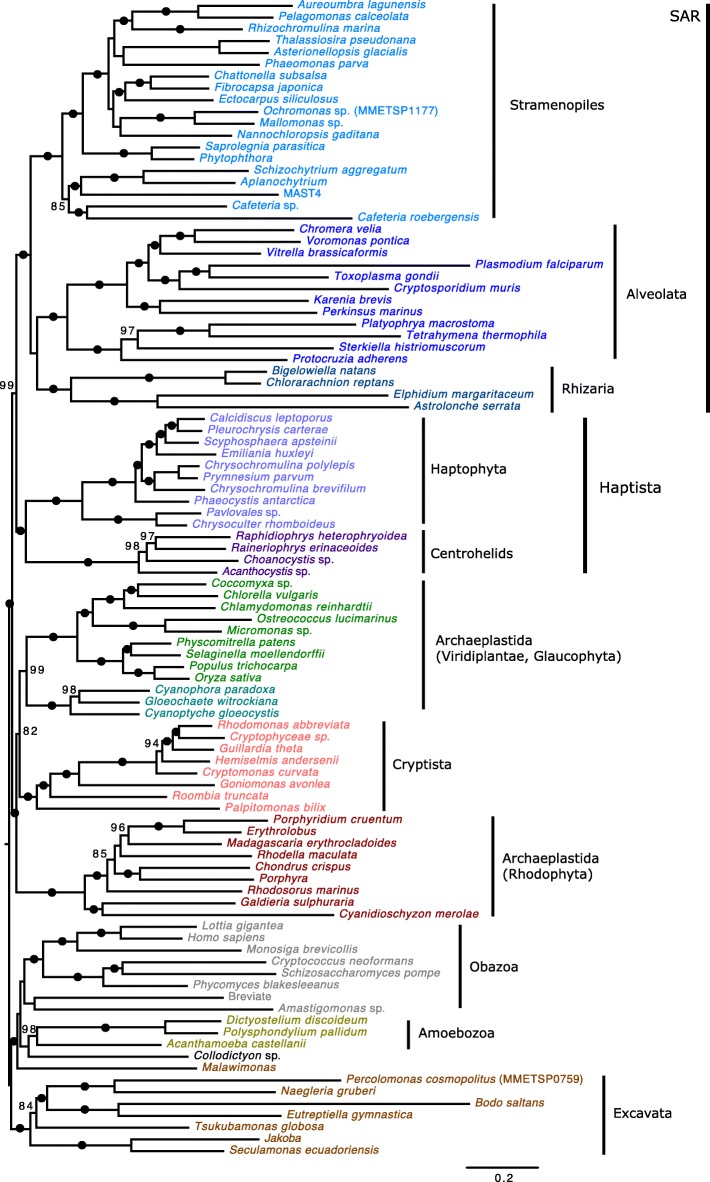
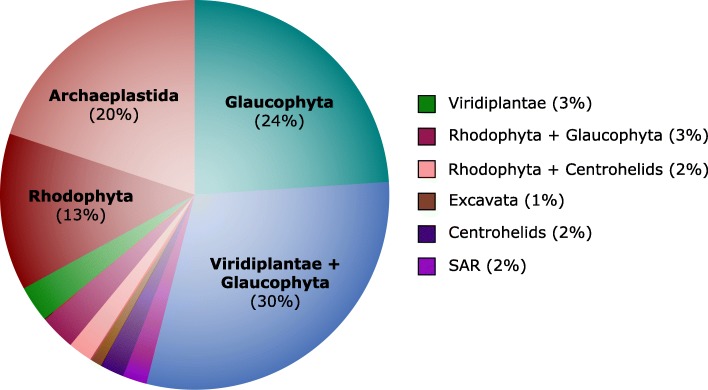
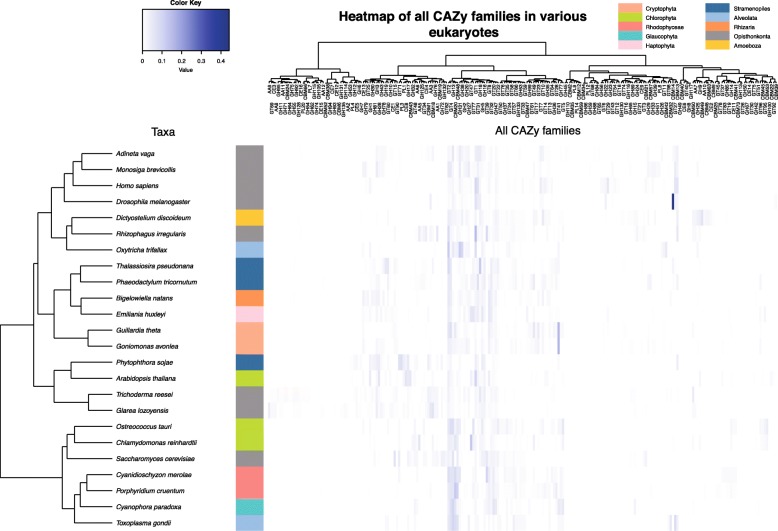
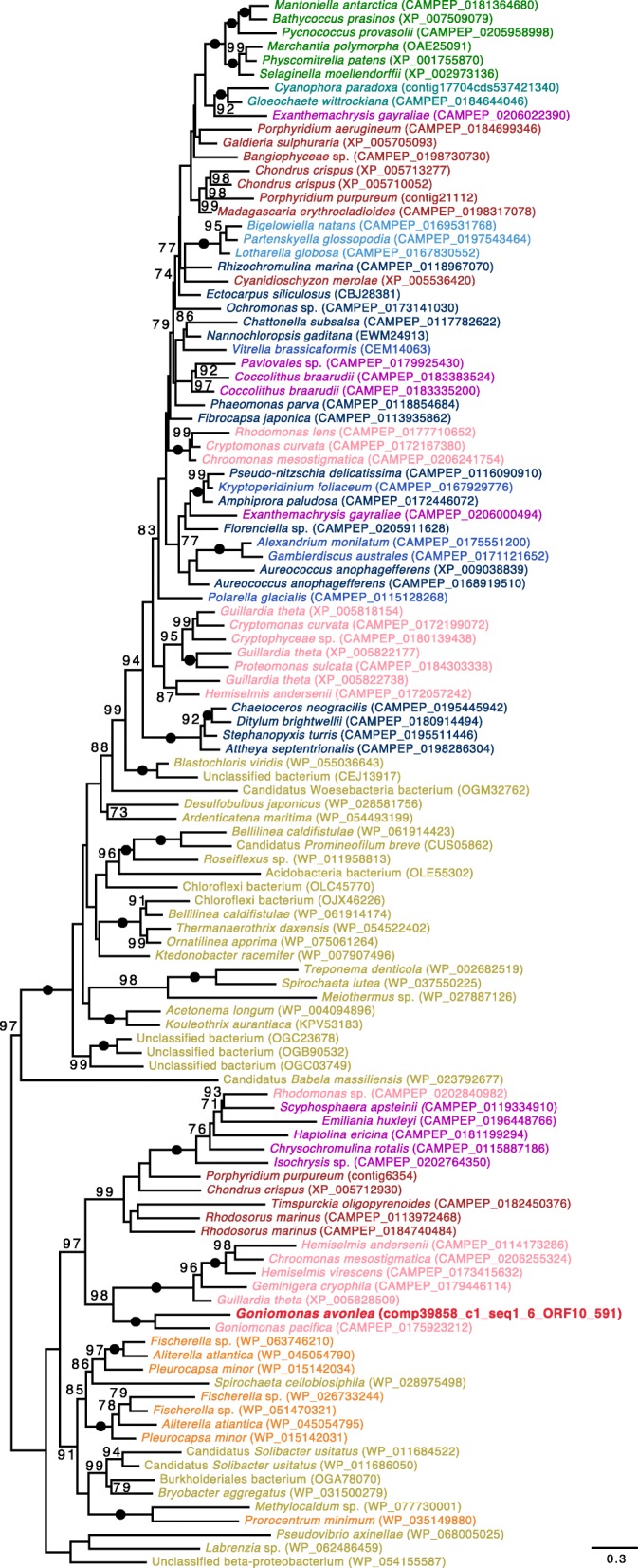
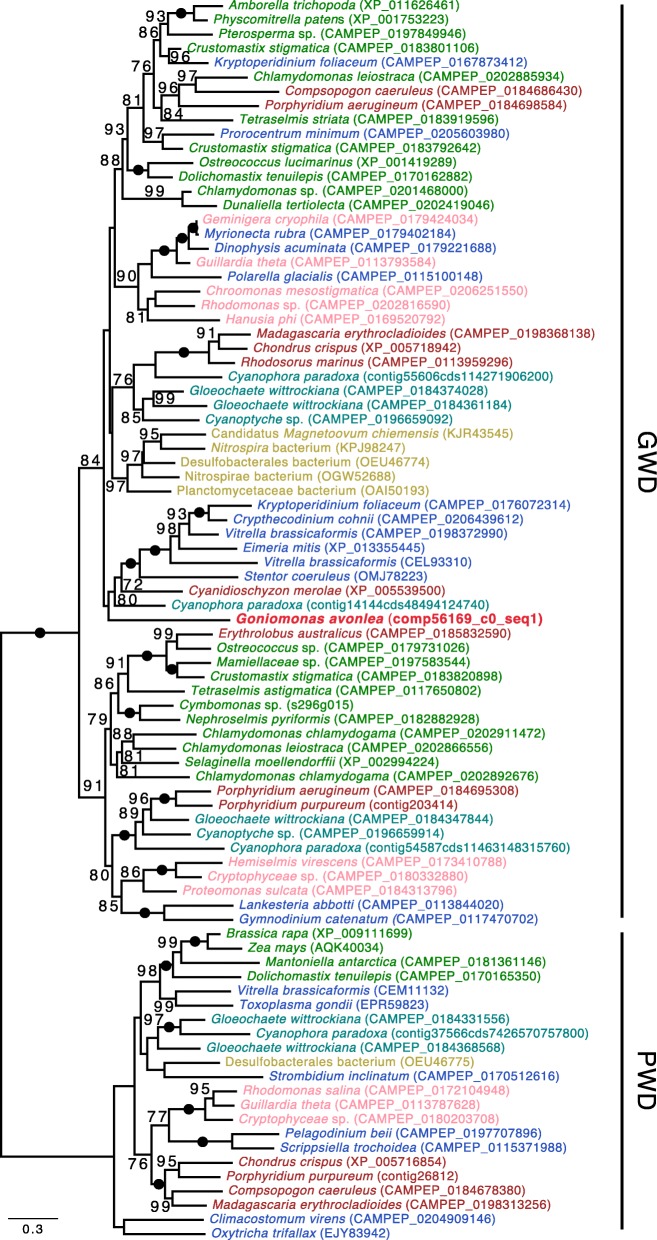
Results: We sequenced the nuclear and mitochondrial genomes of Goniomonas avonlea and carried out a comparative analysis of Go. avonlea, Gu. theta, and other cryptomonads. The Go. avonlea genome assembly is ~ 92 Mbp in size, with 33,470 predicted protein-coding genes. Interestingly, some metabolic pathways (e.g., fatty acid biosynthesis) predicted to occur in the plastid and periplastidal compartment of Gu. theta appear to operate in the cytoplasm of Go. avonlea, suggesting that metabolic redundancies were generated during the course of secondary plastid integration. Other cytosolic pathways found in Go. avonlea are not found in Gu. theta, suggesting secondary loss in Gu. theta and other plastid-bearing cryptomonads. Phylogenetic analyses revealed no evidence for algal endosymbiont-derived genes in the Go. avonlea genome. Phylogenomic analyses point to a specific relationship between Cryptista (to which cryptomonads belong) and Archaeplastida.
Conclusion: We found no convincing genomic or phylogenomic evidence that Go. avonlea evolved from a secondary red algal plastid-bearing ancestor, consistent with goniomonads being ancestrally non-photosynthetic eukaryotes. The Go. avonlea genome sheds light on the physiology of heterotrophic cryptomonads and serves as an important reference point for studying the metabolic "rewiring" that took place during secondary plastid integration in the ancestor of modern-day Cryptophyceae.
Keywords: Cryptomonads; Cryptophytes; Genome evolution; Phylogenomics; Secondary endosymbiosis.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Comment in
-
Genome of tiny predator with big appetite.BMC Biol. 2018 Nov 28;16(1):140. doi: 10.1186/s12915-018-0610-8. BMC Biol. 2018. PMID: 30482182 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
