

The chicken gut metagenome and the modulatory effects of plant-derived benzylisoquinoline alkaloids
- PMID: 30482240
- PMCID: PMC6260706
- DOI: 10.1186/s40168-018-0590-5
The chicken gut metagenome and the modulatory effects of plant-derived benzylisoquinoline alkaloids
Abstract
Background: Sub-therapeutic antibiotics are widely used as growth promoters in the poultry industry; however, the resulting antibiotic resistance threatens public health. A plant-derived growth promoter, Macleaya cordata extract (MCE), with effective ingredients of benzylisoquinoline alkaloids, is a potential alternative to antibiotic growth promoters. Altered intestinal microbiota play important roles in growth promotion, but the underlying mechanism remains unknown.
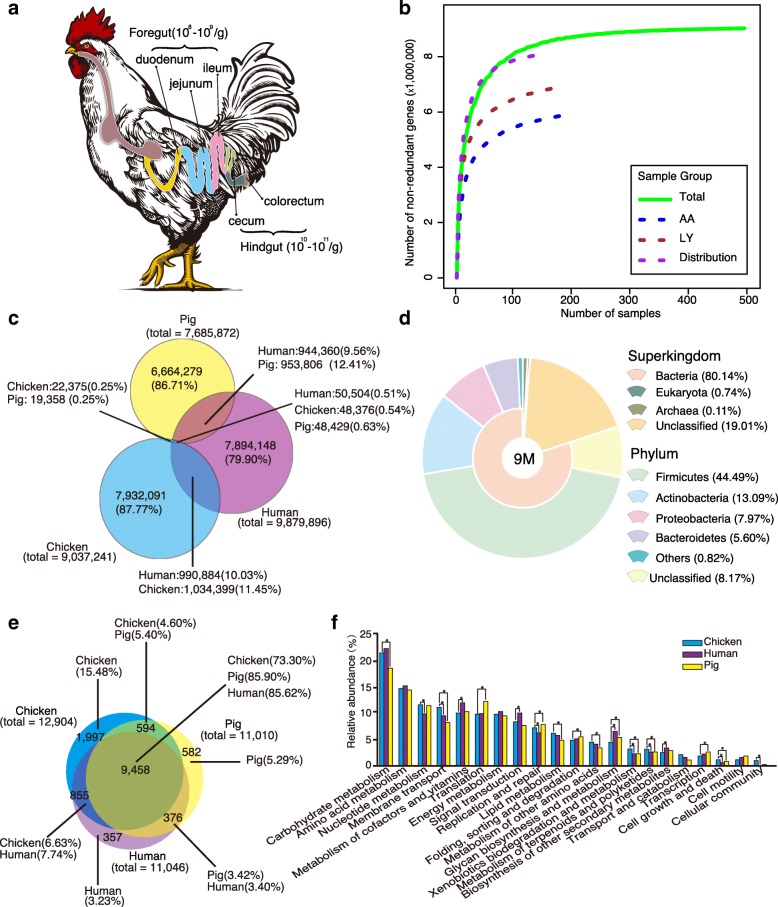
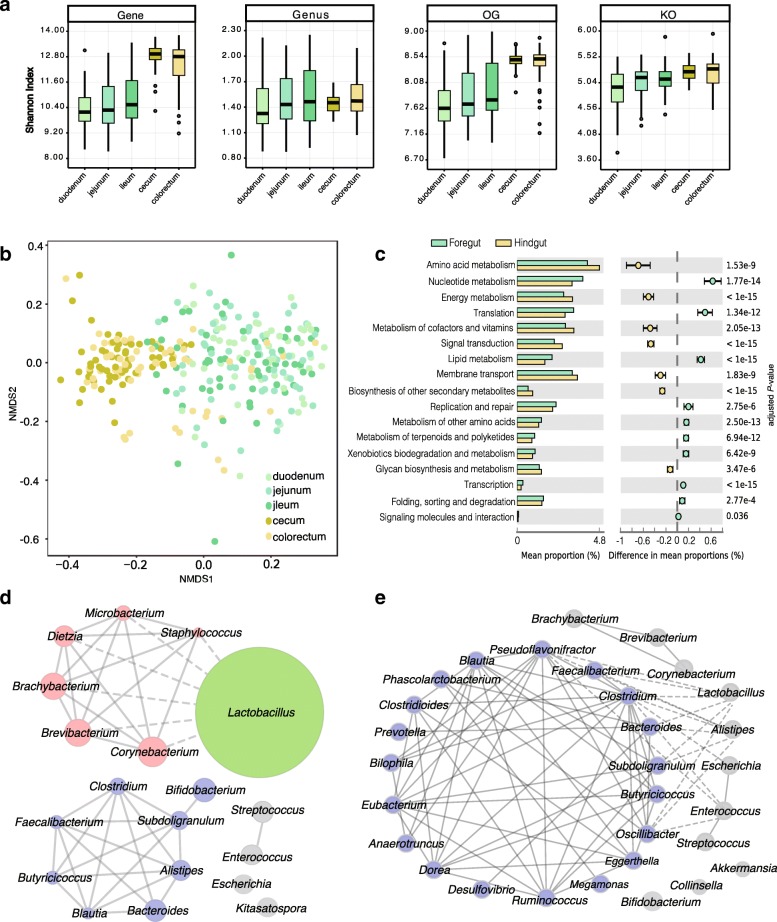
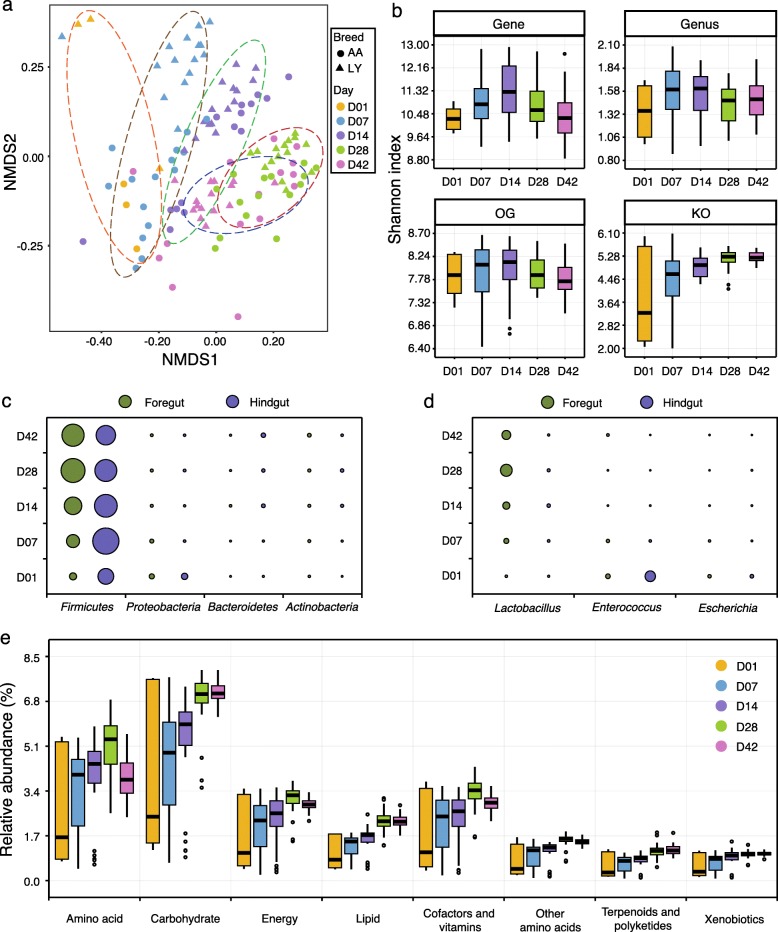
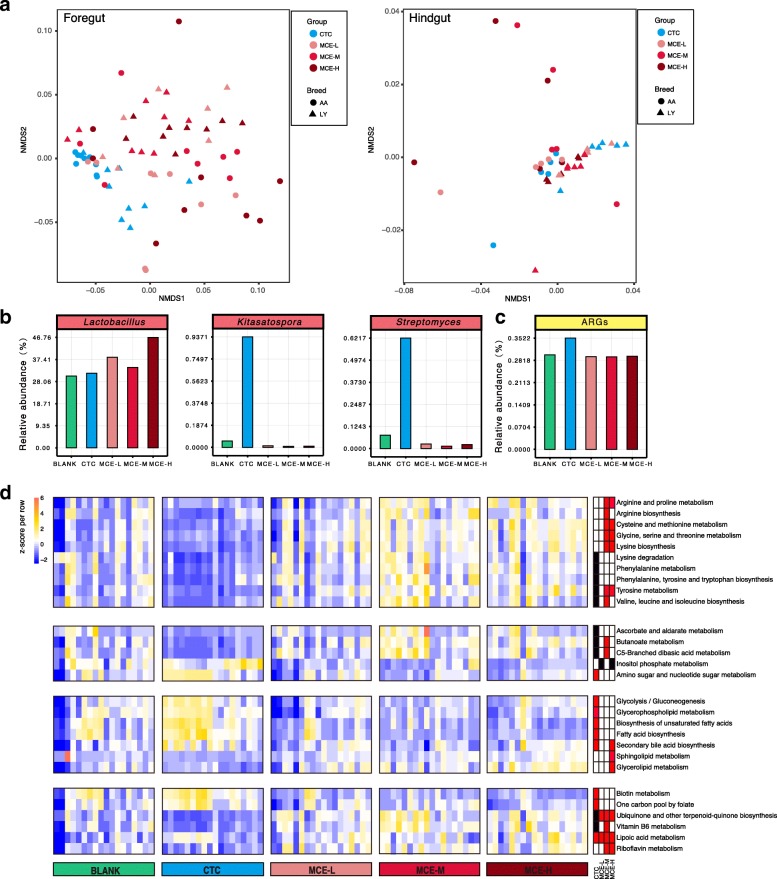
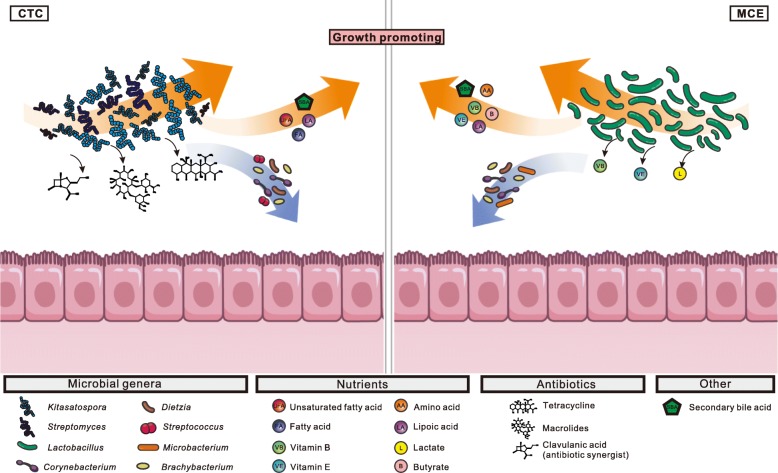
Results: We generated 1.64 terabases of metagenomic data from 495 chicken intestinal digesta samples and constructed a comprehensive chicken gut microbial gene catalog (9.04 million genes), which is also the first gene catalog of an animal's gut microbiome that covers all intestinal compartments. Then, we identified the distinctive characteristics and temporal changes in the foregut and hindgut microbiota. Next, we assessed the impact of MCE on chickens and gut microbiota. Chickens fed with MCE had improved growth performance, and major microbial changes were confined to the foregut, with the predominant role of Lactobacillus being enhanced, and the amino acids, vitamins, and secondary bile acids biosynthesis pathways being upregulated, but lacked the accumulation of antibiotic-resistance genes. In comparison, treatment with chlortetracycline similarly enriched some biosynthesis pathways of nutrients in the foregut microbiota, but elicited an increase in antibiotic-producing bacteria and antibiotic-resistance genes.
Conclusion: The reference gene catalog of the chicken gut microbiome is an important supplement to animal gut metagenomes. Metagenomic analysis provides insights into the growth-promoting mechanism of MCE, and underscored the importance of utilizing safe and effective growth promoters.
Keywords: Antibiotic; Benzylisoquinoline alkaloid; Chicken; Chlortetracycline; Growth promoter; Gut metagenome; Microbiome.
Conflict of interest statement
Ethics approval
This study was approved by the China Agricultural University Animal Care and Use Committee (CAU 20150428-2).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance. Geneva: World Health Organization; 2014.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
