

Aspirin ameliorates experimental autoimmune encephalomyelitis through interleukin-11-mediated protection of regulatory T cells
- PMID: 30482850
- PMCID: PMC6325078
- DOI: 10.1126/scisignal.aar8278
Aspirin ameliorates experimental autoimmune encephalomyelitis through interleukin-11-mediated protection of regulatory T cells
Abstract
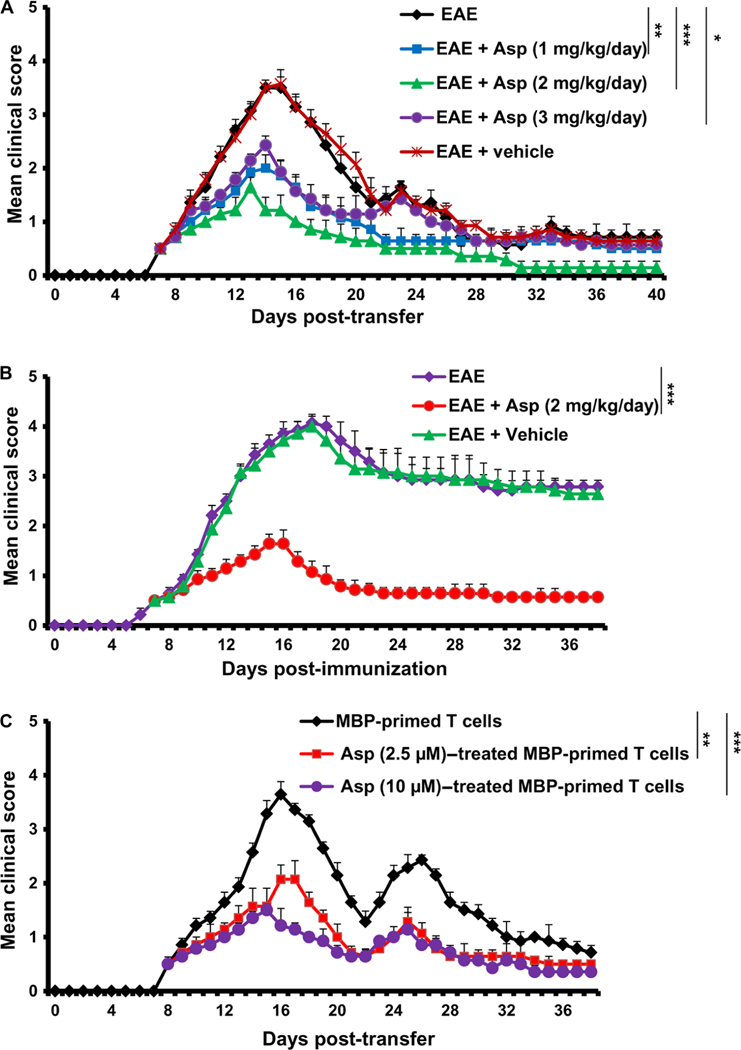
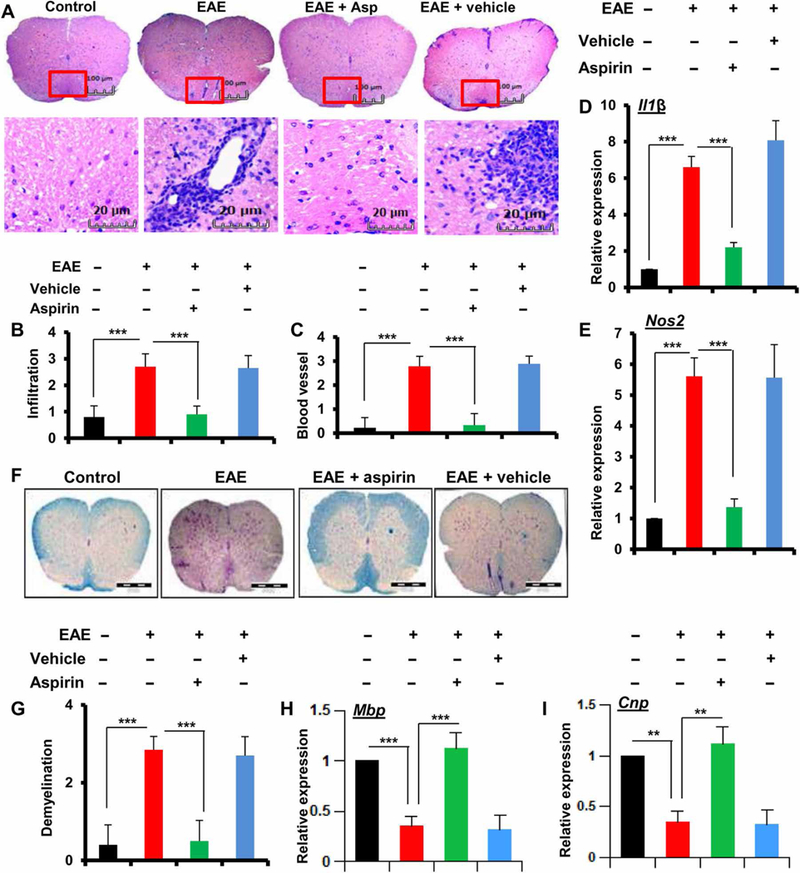
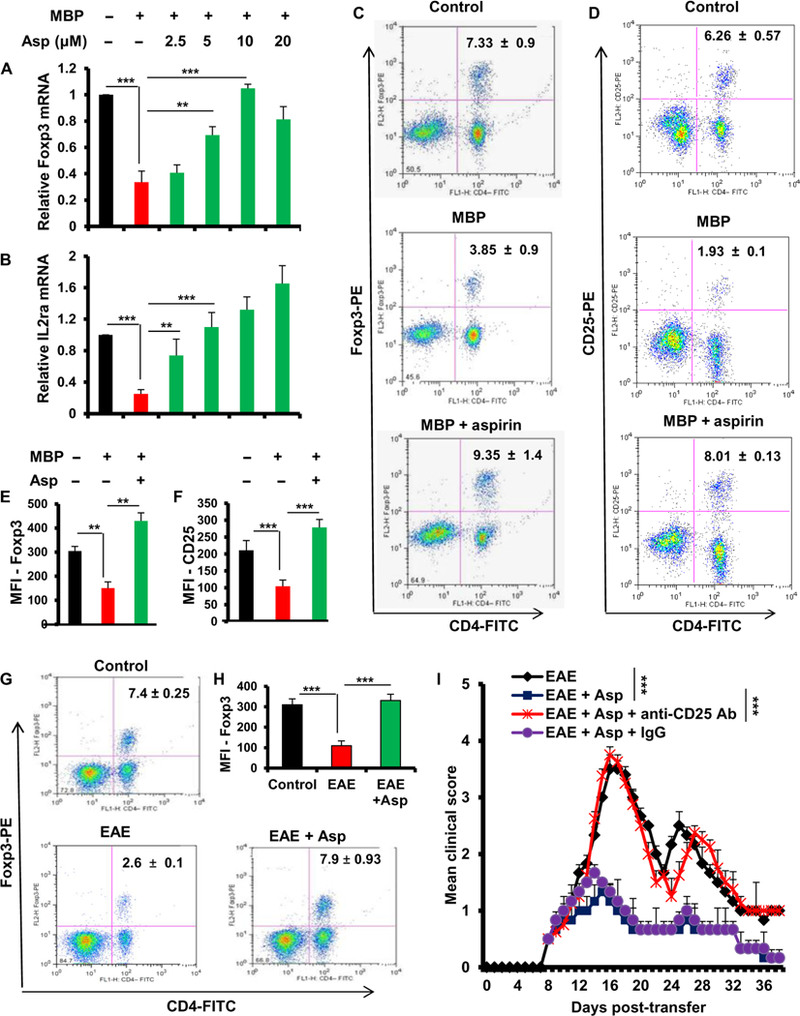
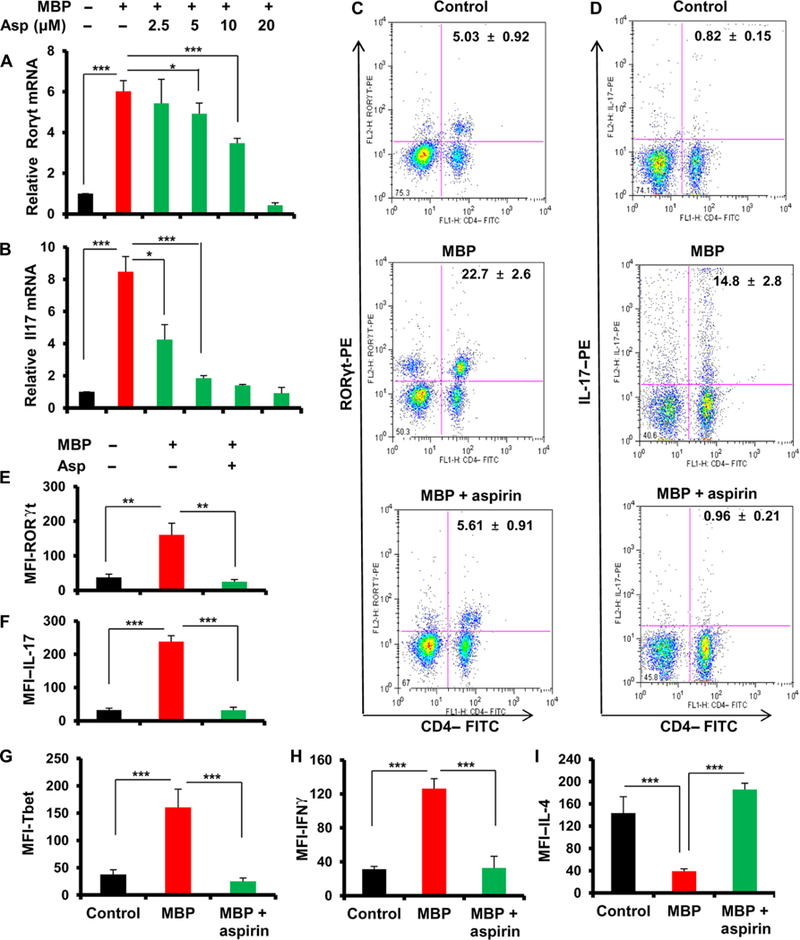
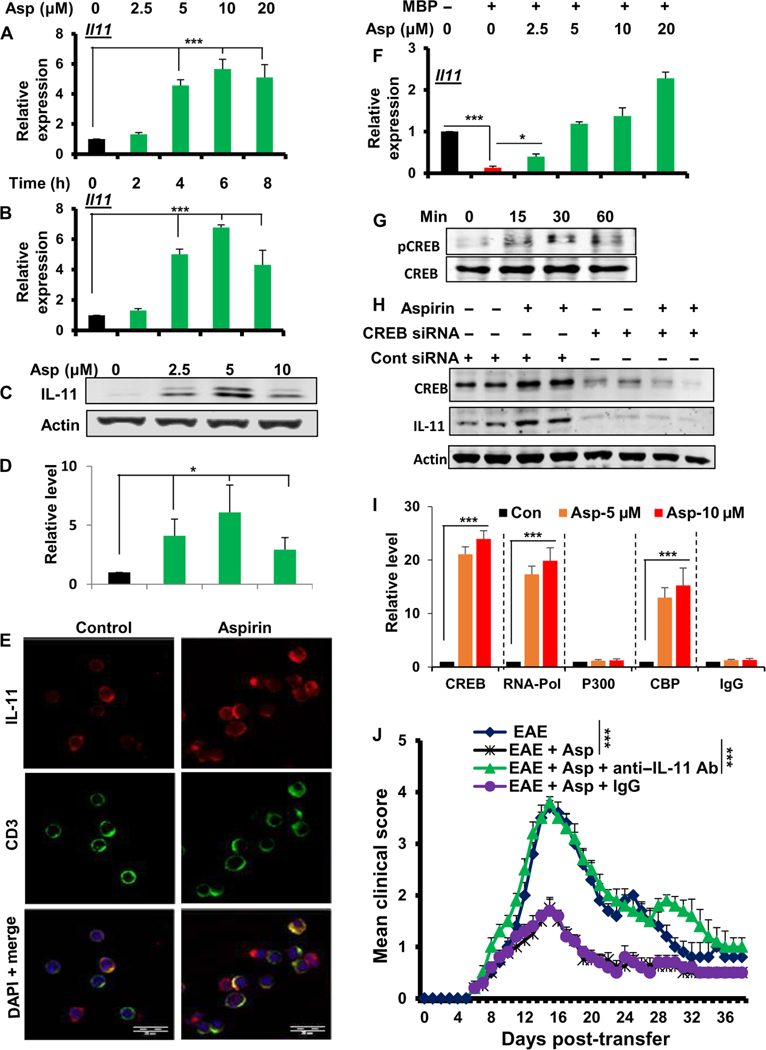
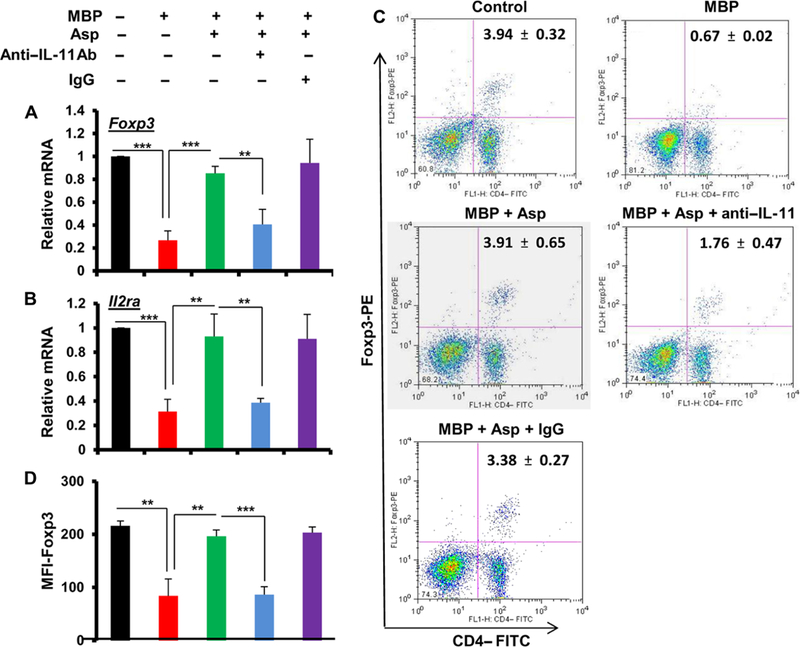
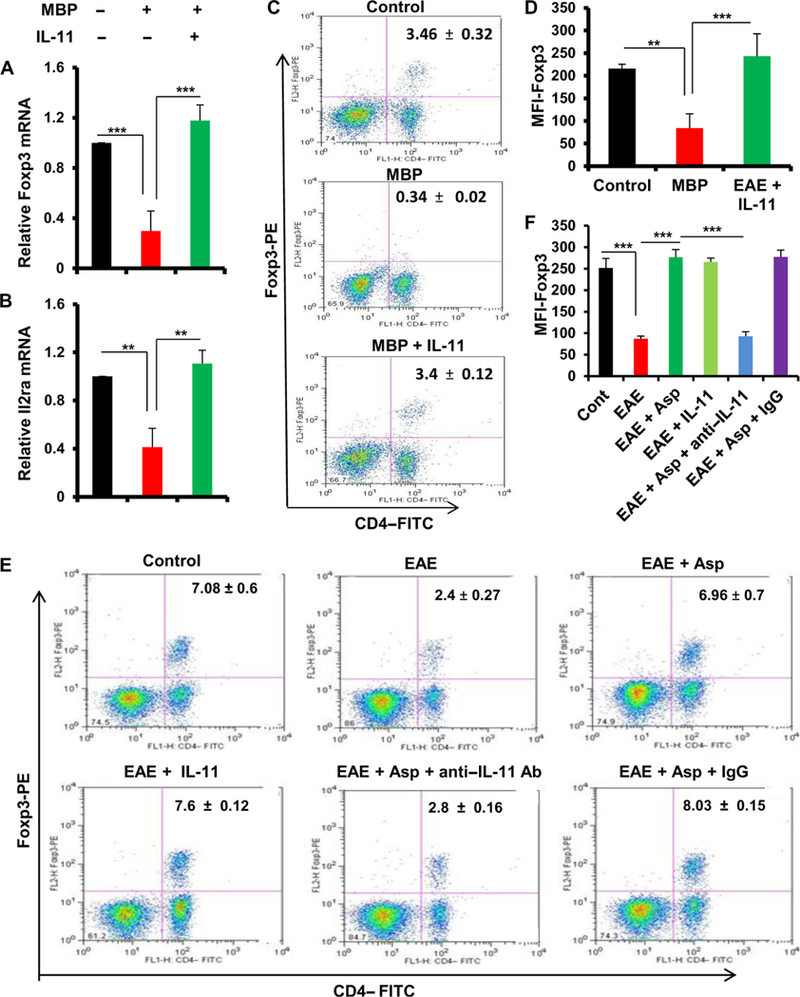
Multiple sclerosis (MS) is a human disease that results from autoimmune T cells targeting myelin protein that is expressed within the central nervous system. In MS, the number of FoxP3-expressing regulatory T cells (Tregs) is reduced, which facilitates the activation of autoreactive T cells. Because aspirin (acetylsalicylic acid) is the most widely used nonsteroidal anti-inflammatory drug, we examined its immunomodulatory effect in mice with experimental autoimmune encephalomyelitis (EAE), an animal model of MS. We found that low-dose aspirin suppressed the clinical symptoms of EAE in mouse models of both relapsing-remitting and chronic disease. Aspirin reduced the development of EAE driven by myelin basic protein (MBP)-specific T cells and the associated perivascular cuffing, inflammation, and demyelination. The effects of aspirin required the presence of CD25+FoxP3+ Tregs Aspirin increased the amounts of Foxp3 and interleukin-4 (IL-4) in T cells and suppressed the differentiation of naïve T cells into T helper 17 (TH17) and TH1 cells. Aspirin also increased the transcription of Il11 mediated by the transcription factor CREB, which was necessary for the generation of Tregs Neutralization of IL-11 negated the effects of aspirin on Treg development and exacerbated EAE. Furthermore, we found that IL-11 alone was sufficient to maintain the percentage of FoxP3+ Tregs and protect mice from EAE. These results identify a previously uncharacterized mode of action of aspirin.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Martin R, McFarland H F, McFarlin D E, Immunological aspects of demyelinating diseases. Annu. Rev. Immunol 10, 153–187 (1992). - PubMed
-
- Coffer P J, Burgering B M T , Forkhead-box transcription factors and their role in the immune system. Nat. Rev. Immunol 4, 889–899 (2004). - PubMed
-
- Sakaguchi S, Powrie F, Emerging challenges in regulatory T cell function and biology. Science 317, 627–629 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
