

Selective Autophagy and Xenophagy in Infection and Disease
- PMID: 30483501
- PMCID: PMC6243101
- DOI: 10.3389/fcell.2018.00147
Selective Autophagy and Xenophagy in Infection and Disease
Abstract
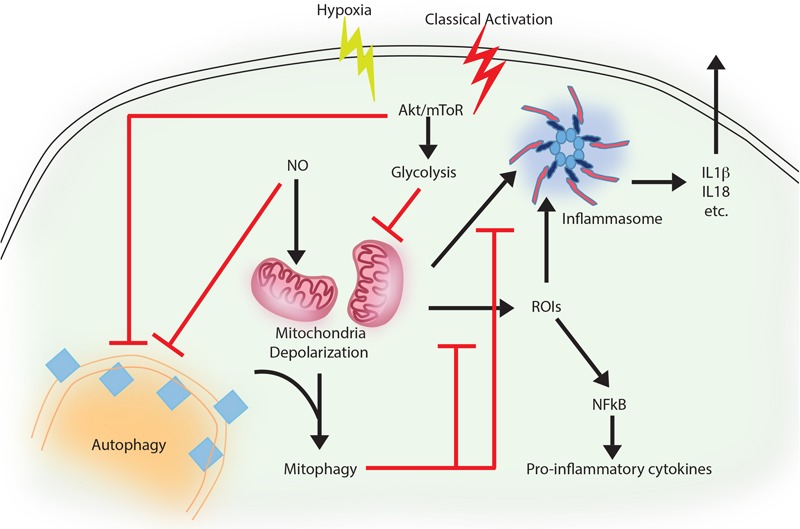
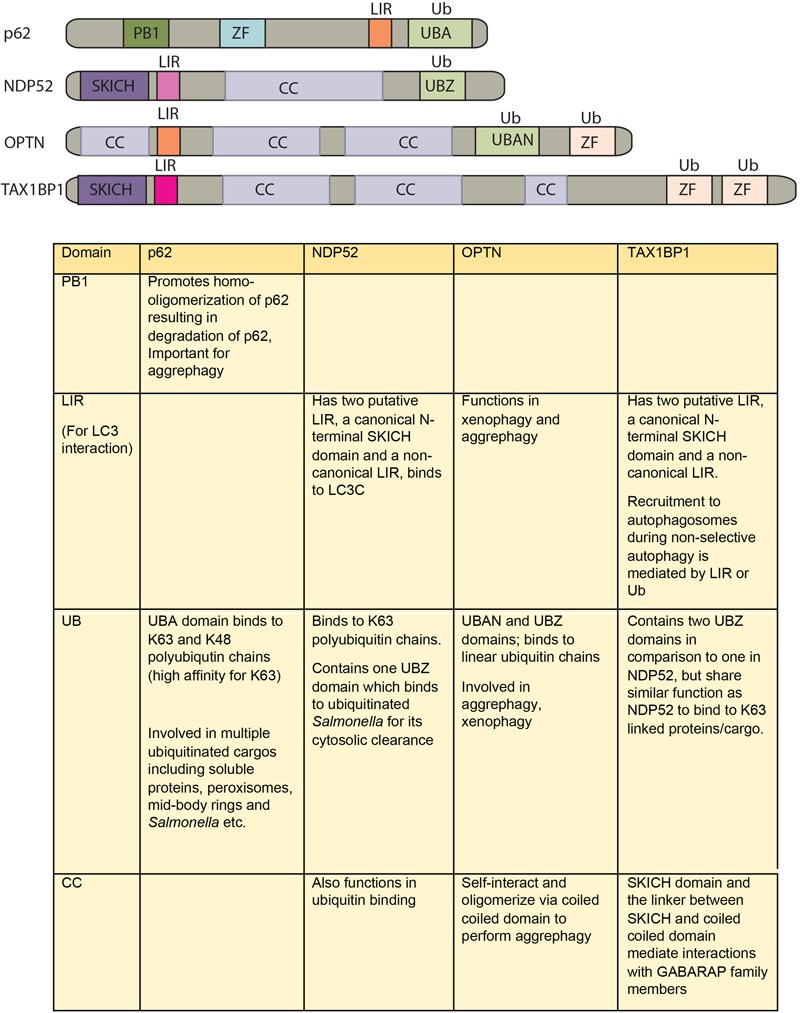
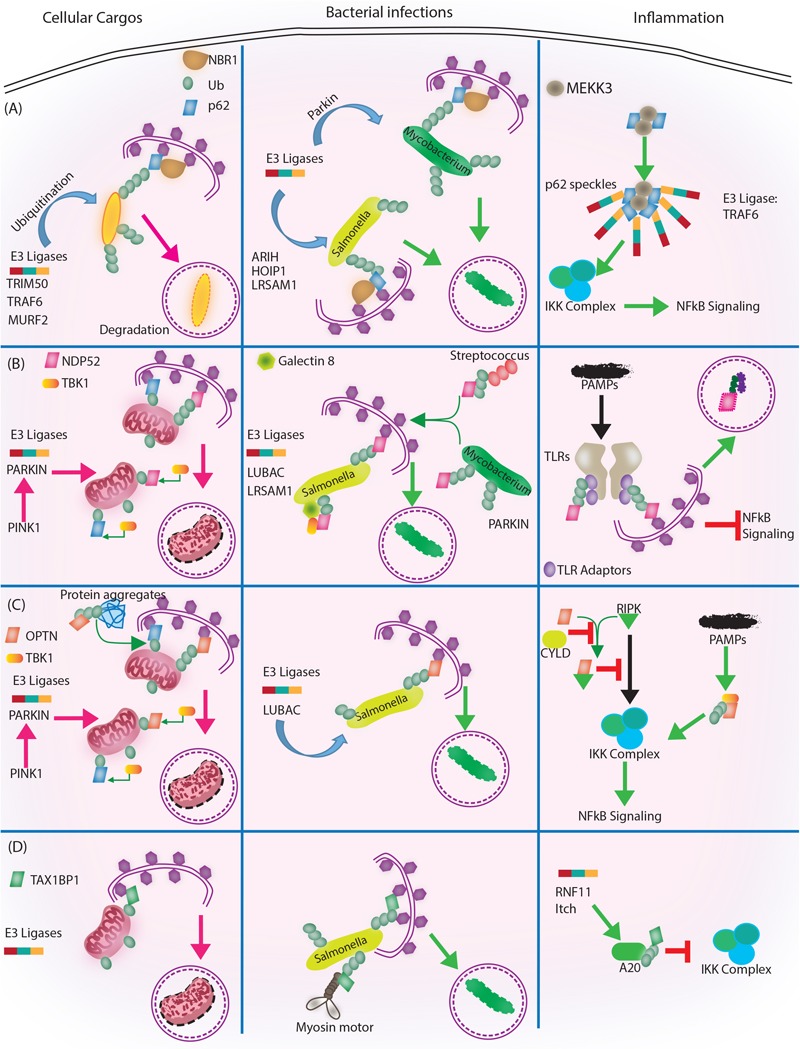
Autophagy, a cellular homeostatic process, which ensures cellular survival under various stress conditions, has catapulted to the forefront of innate defense mechanisms during intracellular infections. The ability of autophagy to tag and target intracellular pathogens toward lysosomal degradation is central to this key defense function. However, studies involving the role and regulation of autophagy during intracellular infections largely tend to ignore the housekeeping function of autophagy. A growing number of evidences now suggest that the housekeeping function of autophagy, rather than the direct pathogen degradation function, may play a decisive role to determine the outcome of infection and immunological balance. We discuss herein the studies that establish the homeostatic and anti-inflammatory function of autophagy, as well as role of bacterial effectors in modulating and coopting these functions. Given that the core autophagy machinery remains largely the same across diverse cargos, how selectivity plays out during intracellular infection remains intriguing. We explore here, the contrasting role of autophagy adaptors being both selective as well as pleotropic in functions and discuss whether E3 ligases could bring in the specificity to cargo selectivity.
Keywords: DUBs; NDP52; OPTN; TAX1BP1; inflammation; p62; ubiquitination; xenophagy.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous