

Multilayered regulation of TORC1-body formation in budding yeast
- PMID: 30485160
- PMCID: PMC6589571
- DOI: 10.1091/mbc.E18-05-0297
Multilayered regulation of TORC1-body formation in budding yeast
Abstract
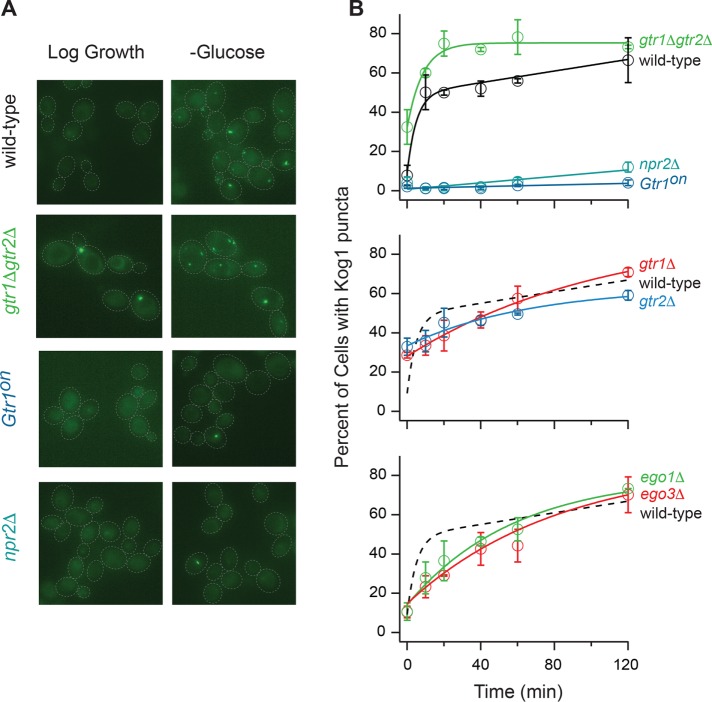
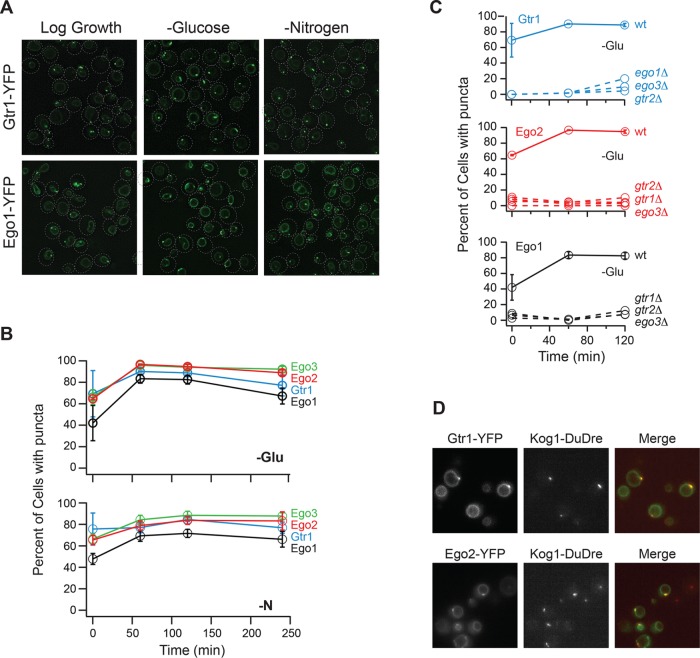
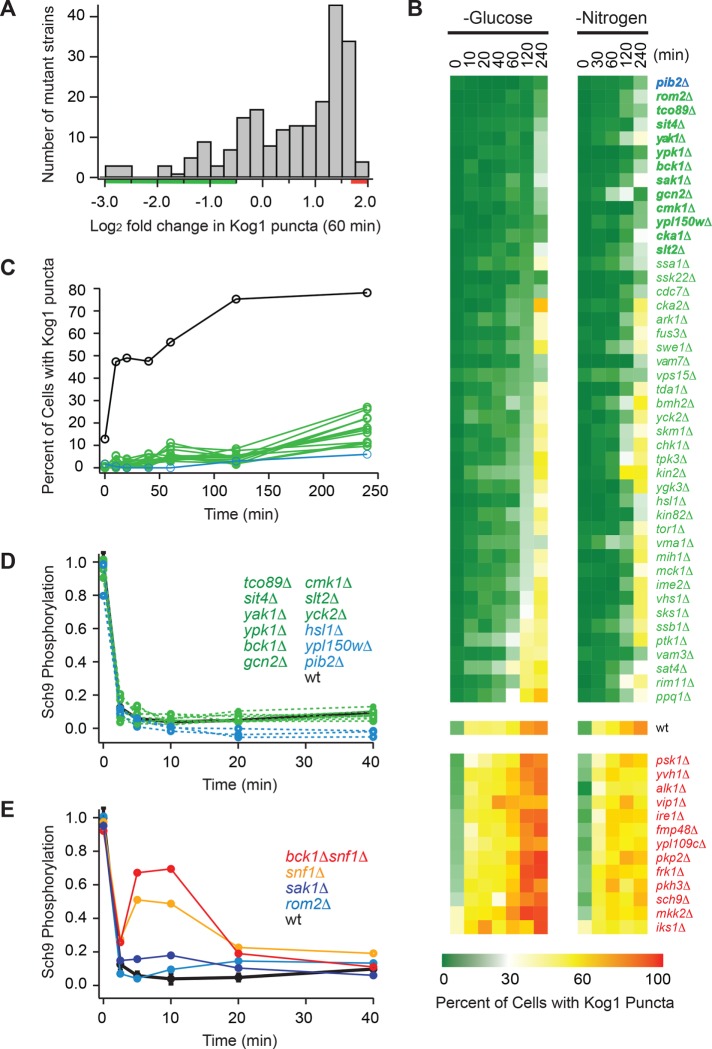
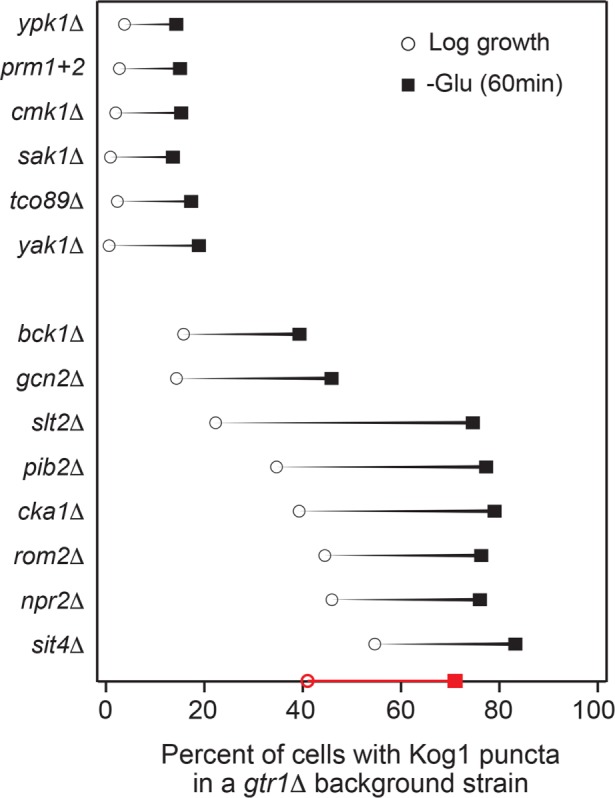
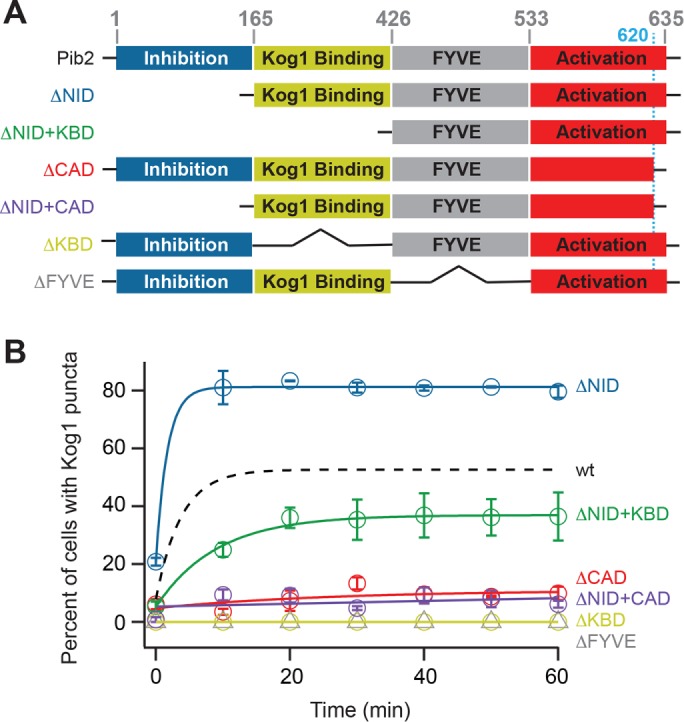
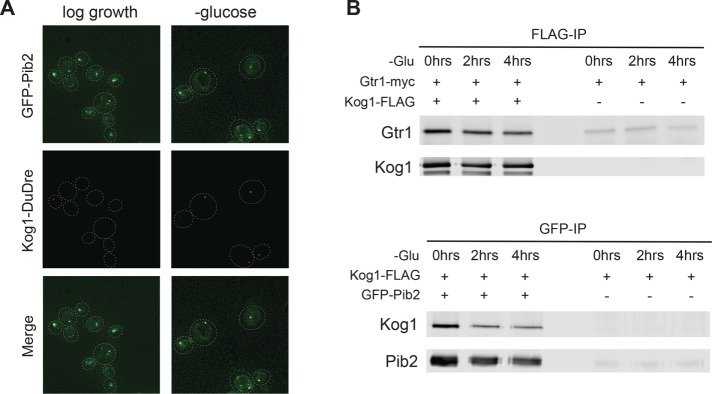
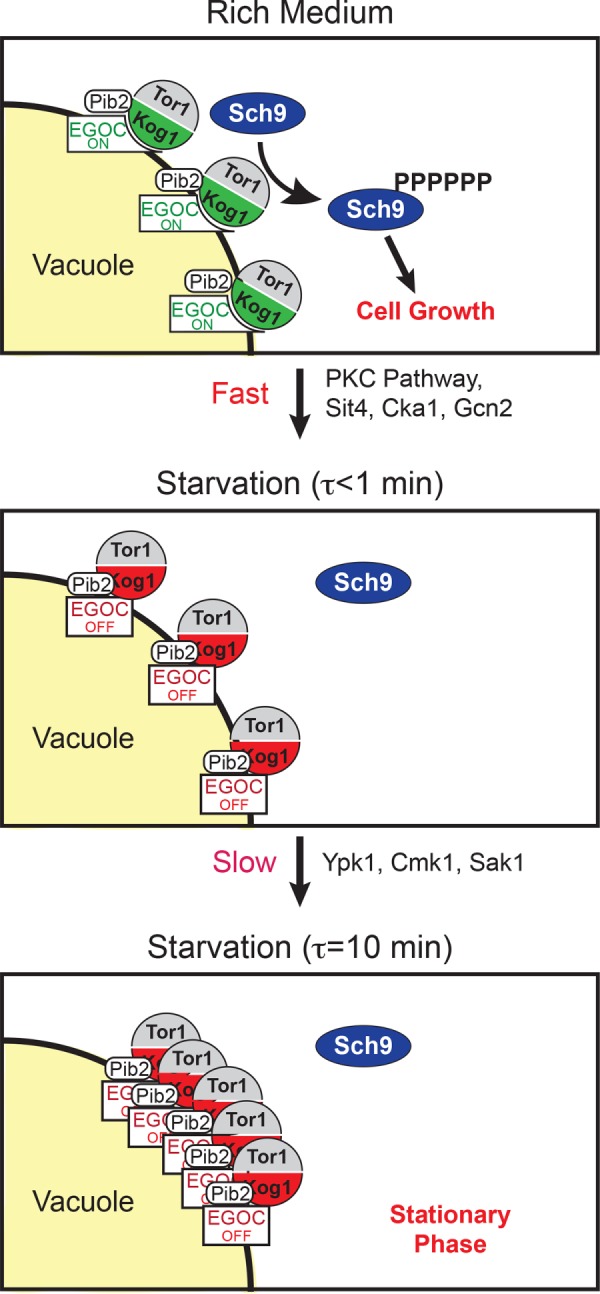
The target of rapamycin kinase complex 1 (TORC1) regulates cell growth and metabolism in eukaryotes. In Saccharomyces cerevisiae, TORC1 activity is known to be controlled by the conserved GTPases, Gtr1/2, and movement into and out of an inactive agglomerate/body. However, it is unclear whether/how these regulatory steps are coupled. Here we show that active Gtr1/2 is a potent inhibitor of TORC1-body formation, but cells missing Gtr1/2 still form TORC1-bodies in a glucose/nitrogen starvation-dependent manner. We also identify 13 new activators of TORC1-body formation and show that seven of these proteins regulate the Gtr1/2-dependent repression of TORC1-body formation, while the remaining proteins drive the subsequent steps in TORC1 agglomeration. Finally, we show that the conserved phosphatidylinositol-3-phosphate (PI(3)P) binding protein, Pib2, forms a complex with TORC1 and overrides the Gtr1/2-dependent repression of TORC1-body formation during starvation. These data provide a unified, systems-level model of TORC1 regulation in yeast.
Figures
References
-
- Adami A, Garcia-Alvarez B, Arias-Palomo E, Barford D, Llorca O. (2007). Structure of TOR and its complex with KOG1. Mol Cell , 509–516. - PubMed
-
- Aylett CH, Sauer E, Imseng S, Boehringer D, Hall MN, Ban N, Maier T. (2016). Architecture of human mTOR complex 1. Science , 48–52. - PubMed
-
- Binda M, Peli-Gulli MP, Bonfils G, Panchaud N, Urban J, Sturgill TW, Loewith R, De Virgilio C. (2009). The Vam6 GEF controls TORC1 by activating the EGO complex. Mol Cell , 563–573. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
