

Altered expression of K13 disrupts DNA replication and repair in Plasmodium falciparum
- PMID: 30486796
- PMCID: PMC6263542
- DOI: 10.1186/s12864-018-5207-7
Altered expression of K13 disrupts DNA replication and repair in Plasmodium falciparum
Abstract
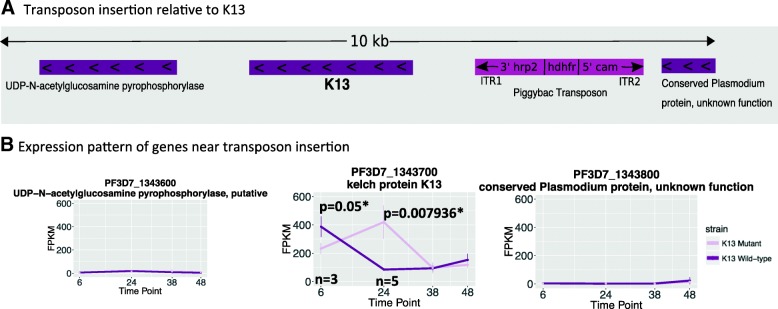
Background: Plasmodium falciparum exhibits resistance to the artemisinin component of the frontline antimalarial treatment Artemisinin-based Combination Therapy in South East Asia. Millions of lives will be at risk if artemisinin resistance (ART-R) spreads to Africa. Single non-synonymous mutations in the propeller region of PF3D7_1343700,"K13" are implicated in resistance. In this work, we use transcriptional profiling to characterize a laboratory-generated k13 insertional mutant previously demonstrated to have increased sensitivity to artemisinins to explore the functional role of k13.
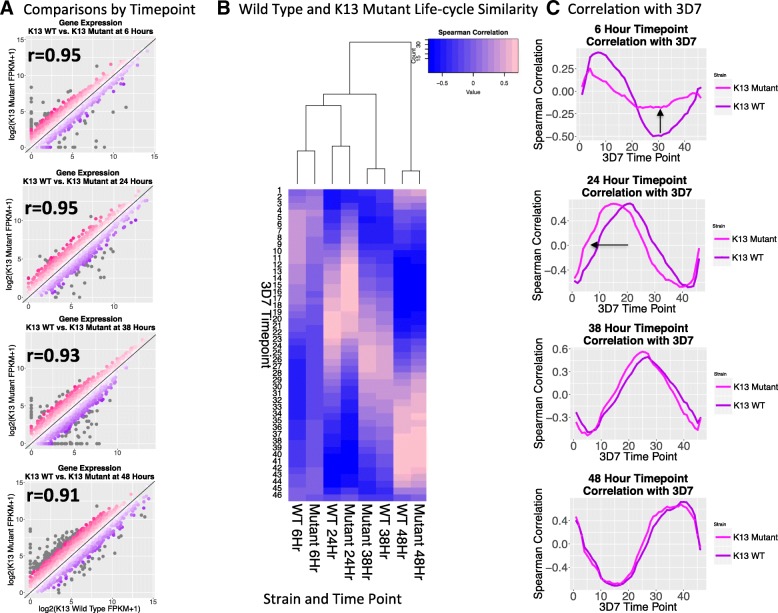
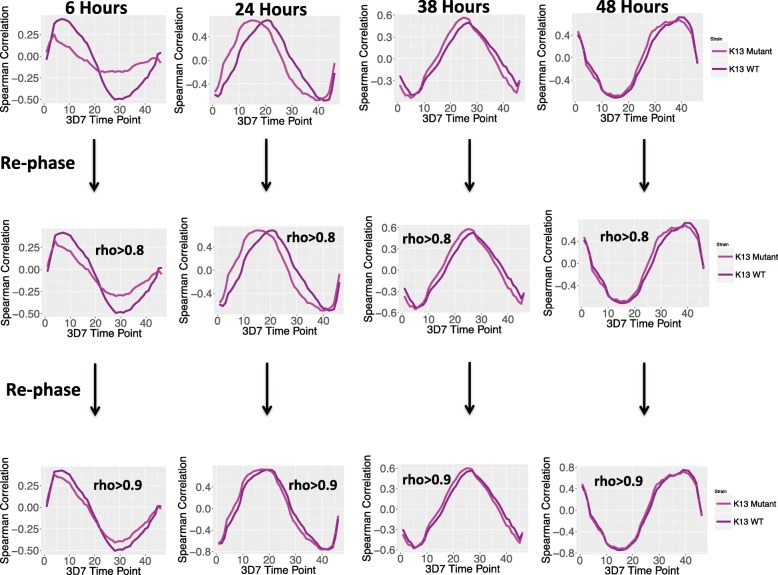
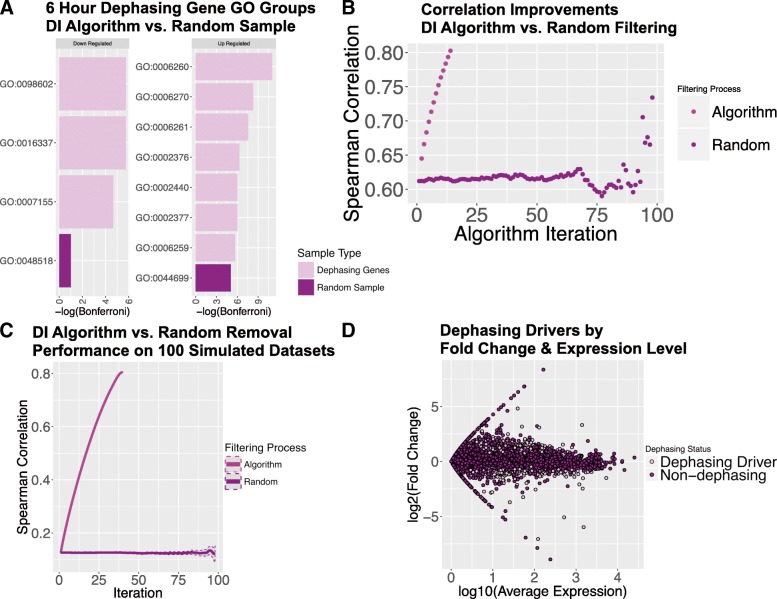
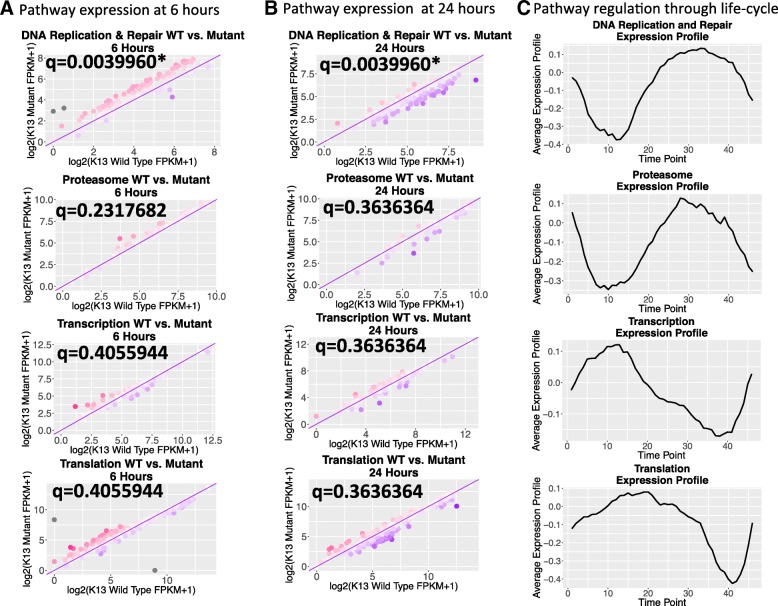
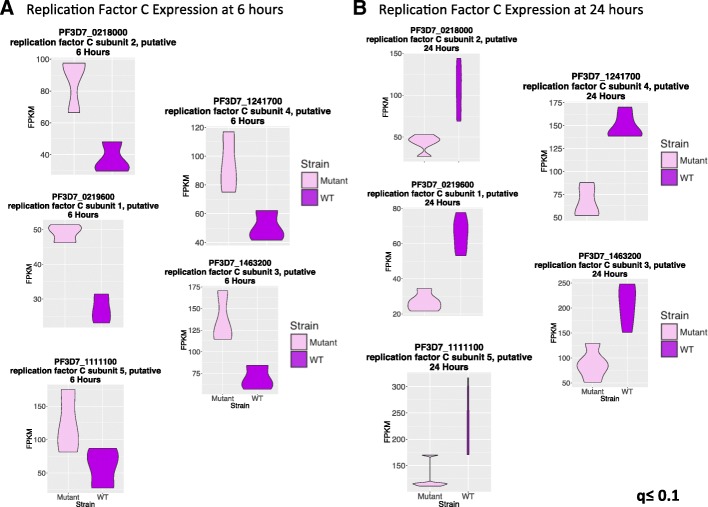
Results: A set of RNA-seq and microarray experiments confirmed that the expression profile of k13 is specifically altered during the early ring and early trophozoite stages of the mutant intraerythrocytic development cycle. The down-regulation of k13 transcripts in this mutant during the early ring stage is associated with a transcriptome advance towards a more trophozoite-like state. To discover the specific downstream effect of k13 dysregulation, we developed a new computational method to search for differential gene expression while accounting for the temporal sequence of transcription. We found that the strongest biological signature of the transcriptome shift is an up-regulation of DNA replication and repair genes during the early ring developmental stage and a down-regulation of DNA replication and repair genes during the early trophozoite stage; by contrast, the expressions of housekeeping genes are unchanged. This effect, due to k13 dysregulation, is antagonistic, such that k13 levels are negatively correlated with DNA replication and repair gene expression.
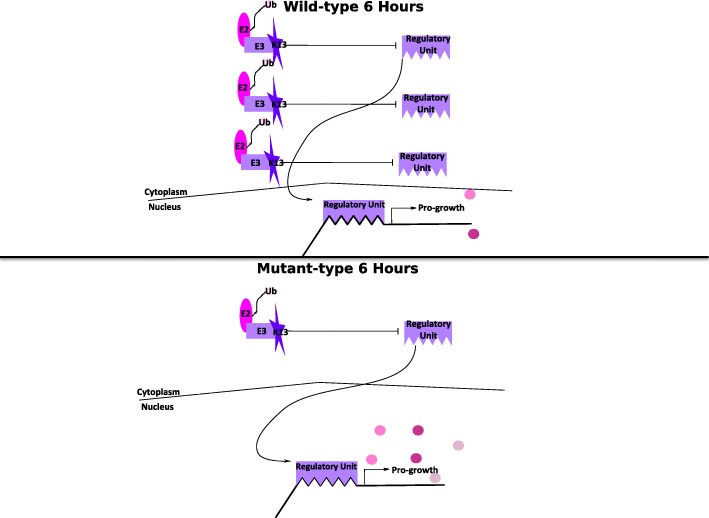
Conclusion: Our results support a role for k13 as a stress response regulator consistent with the hypothesis that artemisinins mode of action is oxidative stress and k13 as a functional homolog of Keap1 which in humans regulates DNA replication and repair genes in response to oxidative stress.
Keywords: Artemisinin; Drug-resistance; K13; Malaria.
Conflict of interest statement
Ethics approval and consent to participate
Ethical approval for the use of human blood in this study was granted by the Institutional Review Boards of the University of South Florida and the University of Notre Dame. All of the blood used for the in vitro culturing of parasites was obtained from healthy adult volunteers and drawn by trained personal from Interstate Blood Bank.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Organization WH . World Malaria Report. 2016. p. 2016.
-
- Organization WH . World Malaria Report. 2005. p. 2005.
-
- Amaratunga Chanaki, Lim Pharath, Suon Seila, Sreng Sokunthea, Mao Sivanna, Sopha Chantha, Sam Baramey, Dek Dalin, Try Vorleak, Amato Roberto, Blessborn Daniel, Song Lijiang, Tullo Gregory S, Fay Michael P, Anderson Jennifer M, Tarning Joel, Fairhurst Rick M. Dihydroartemisinin–piperaquine resistance in Plasmodium falciparum malaria in Cambodia: a multisite prospective cohort study. The Lancet Infectious Diseases. 2016;16(3):357–365. doi: 10.1016/S1473-3099(15)00487-9. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
