

Two functionally redundant sources of retinoic acid secure spermatogonia differentiation in the seminiferous epithelium
- PMID: 30487180
- PMCID: PMC6340151
- DOI: 10.1242/dev.170225
Two functionally redundant sources of retinoic acid secure spermatogonia differentiation in the seminiferous epithelium
Abstract
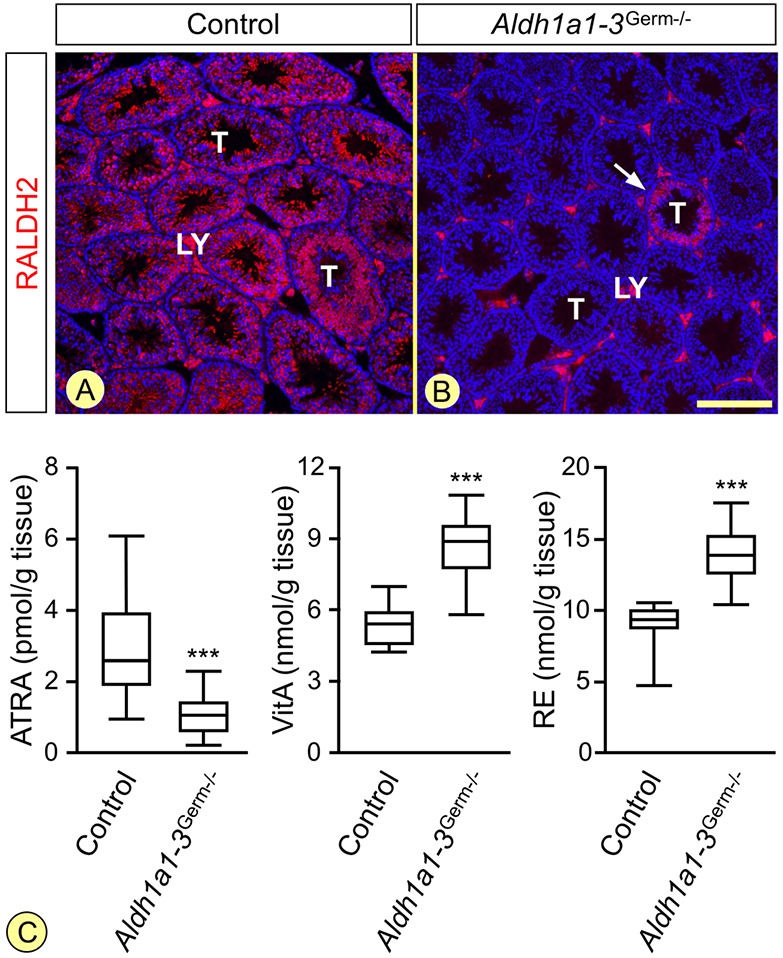
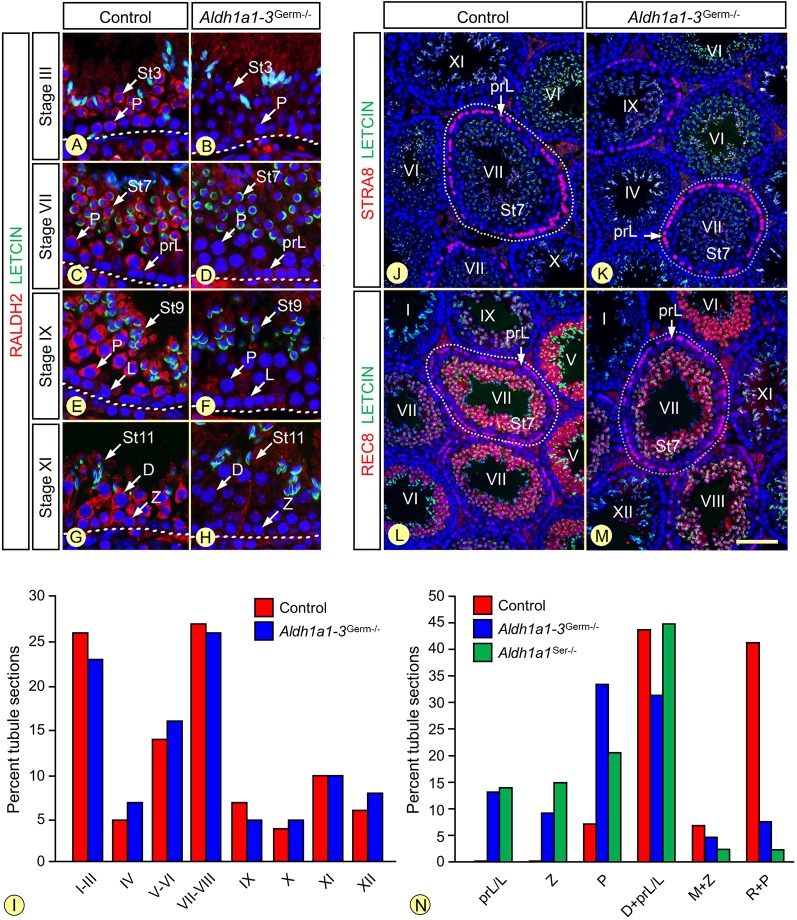
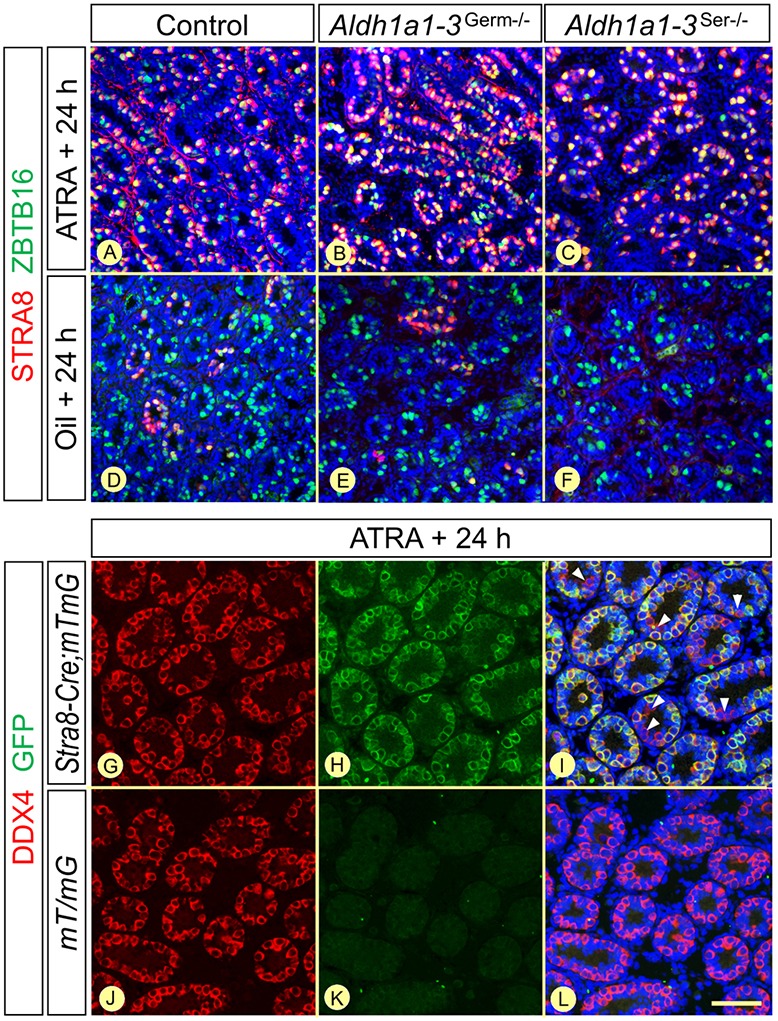
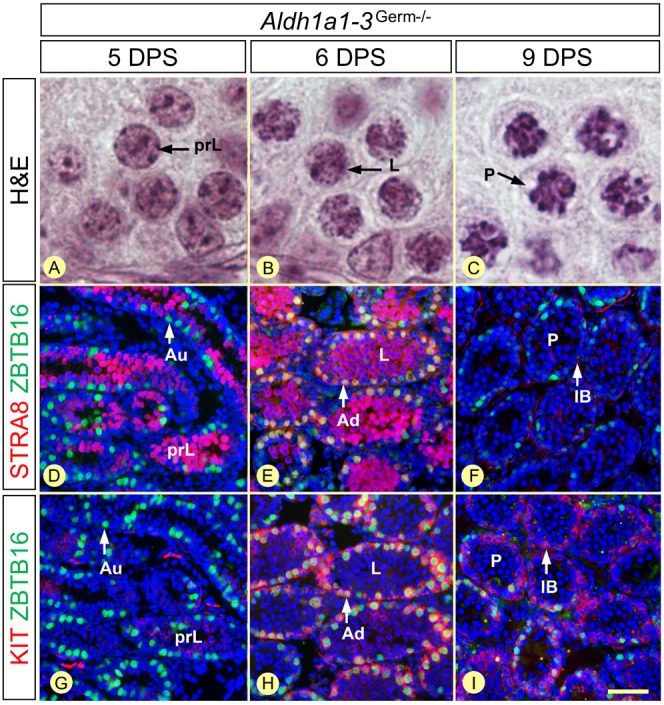
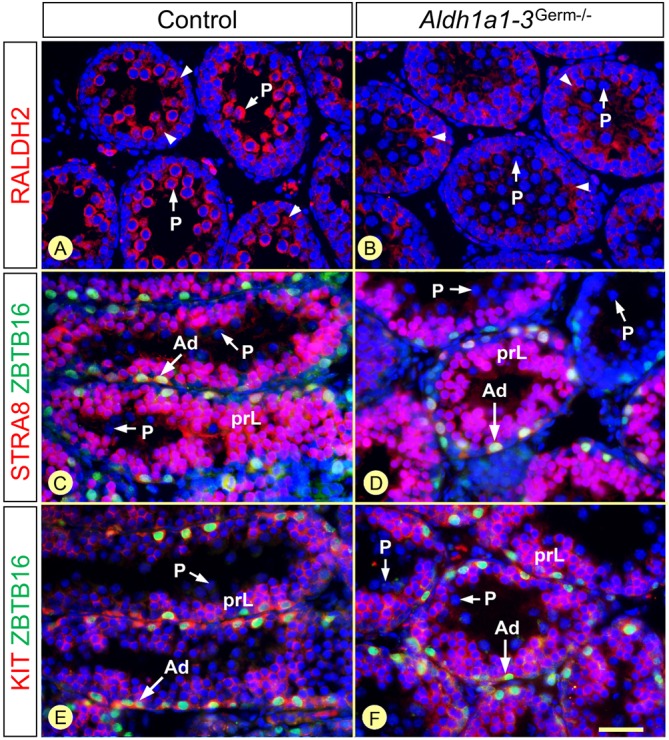
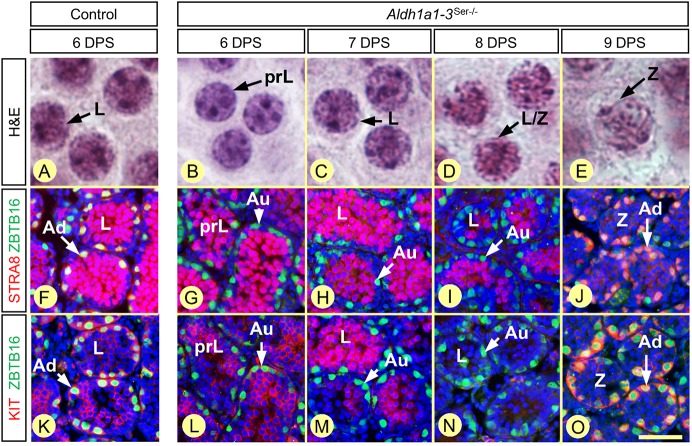
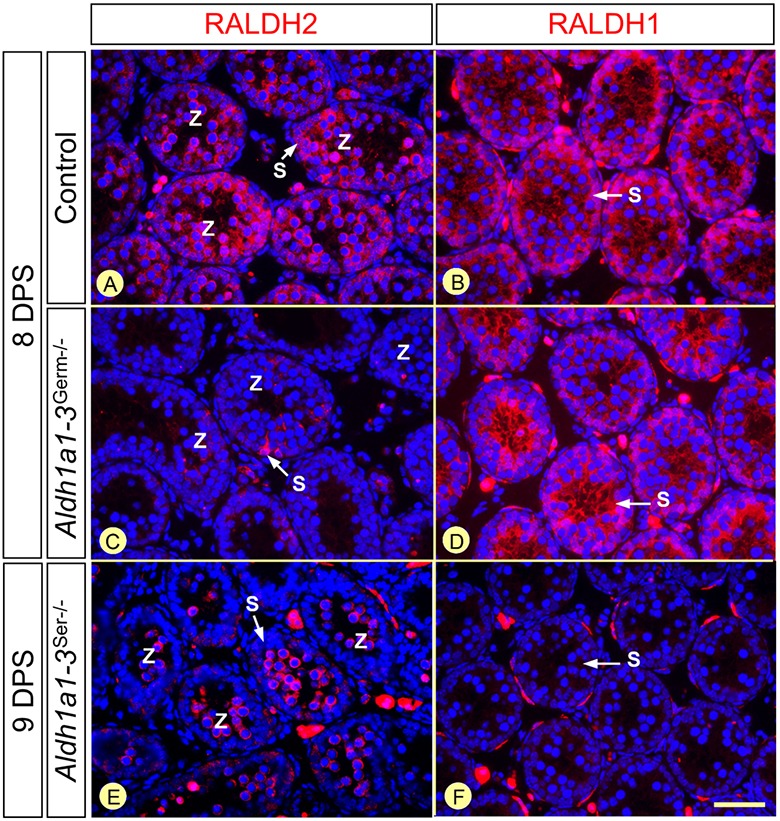
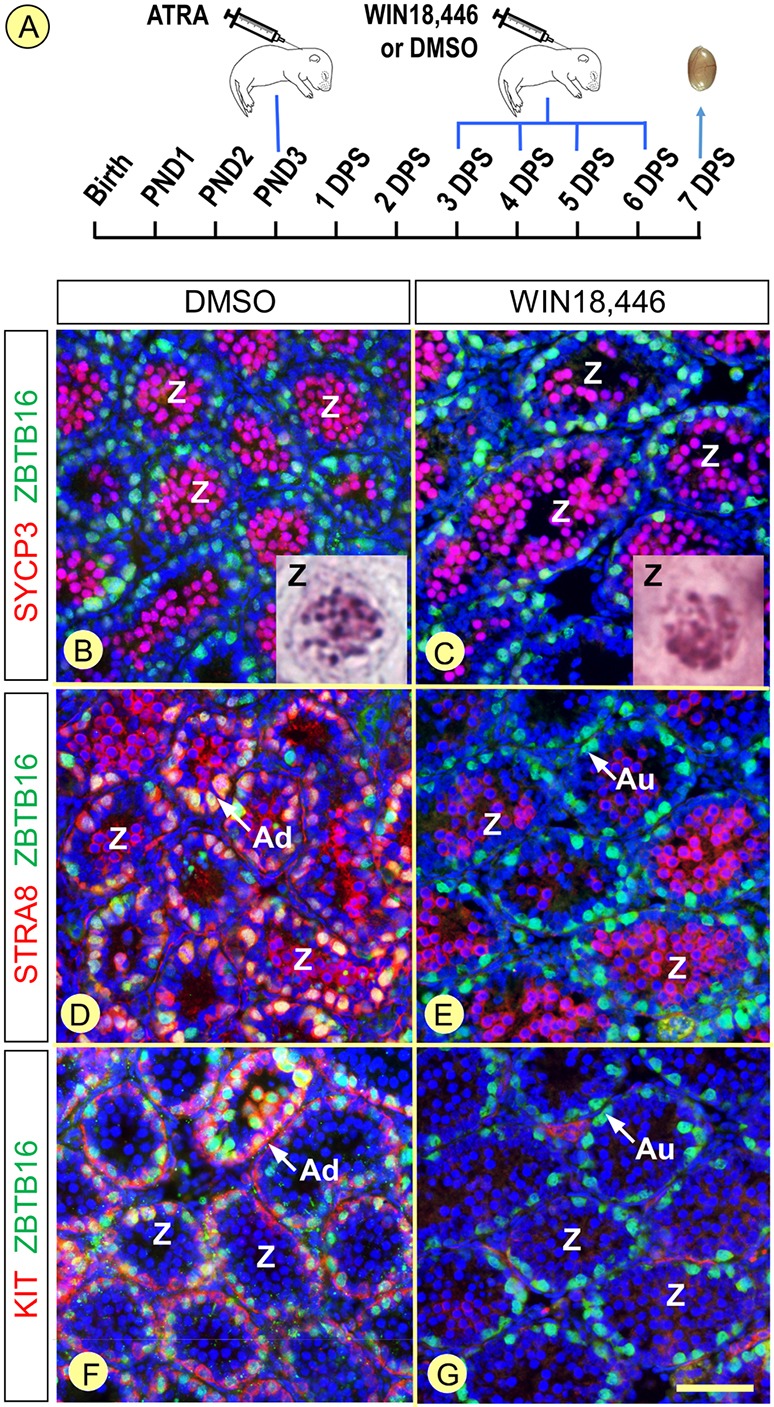
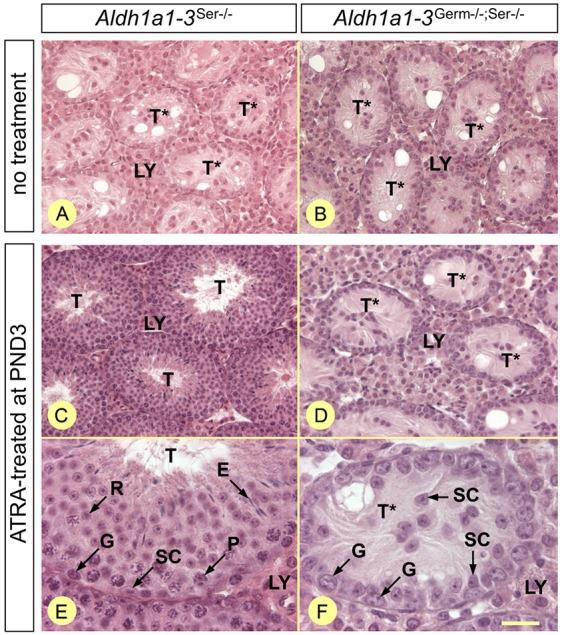
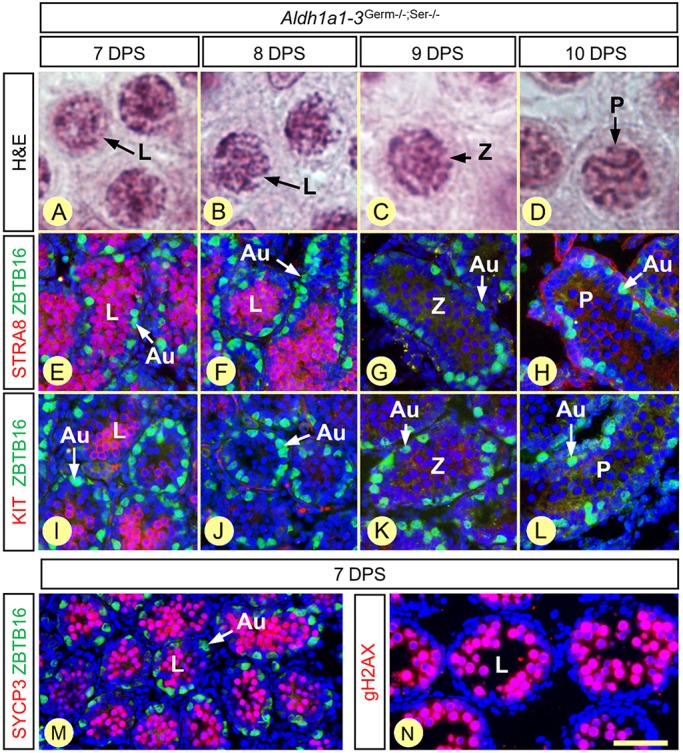
In mammals, all-trans retinoic acid (ATRA) is instrumental to spermatogenesis. It is synthesized by two retinaldehyde dehydrogenases (RALDH) present in both Sertoli cells (SCs) and germ cells (GCs). In order to determine the relative contributions of each source of ATRA, we have generated mice lacking all RALDH activities in the seminiferous epithelium (SE). We show that both the SC- and GC-derived sources of ATRA cooperate to initiate and propagate spermatogenetic waves at puberty. In adults, they exert redundant functions and, against all expectations, the GC-derived source does not perform any specific roles despite contributing to two-thirds of the total amount of ATRA present in the testis. The production from SCs is sufficient to maintain the periodic expression of genes in SCs, as well and the cycle and wave of the SE, which account for the steady production of spermatozoa. The production from SCs is also specifically required for spermiation. Importantly, our study shows that spermatogonia differentiation depends upon the ATRA synthesized by RALDH inside the SE, whereas initiation of meiosis and expression of STRA8 by spermatocytes can occur without ATRA.
Keywords: Aldh8a1 knockout; Immunohistochemistry; Lgals1; Puberty; Spermatogenesis.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Anderson E. L., Baltus A. E., Roepers-Gajadien H. L., Hassold T. J., de Rooij D. G., van Pelt A. M. M. and Page D. C. (2008). Stra8 and its inducer, retinoic acid, regulate meiotic initiation in both spermatogenesis and oogenesis in mice. Proc. Natl. Acad. Sci. USA 105, 14976-14980. 10.1073/pnas.0807297105 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
