

Photoreceptive retinal ganglion cells control the information rate of the optic nerve
- PMID: 30487225
- PMCID: PMC6294960
- DOI: 10.1073/pnas.1810701115
Photoreceptive retinal ganglion cells control the information rate of the optic nerve
Abstract
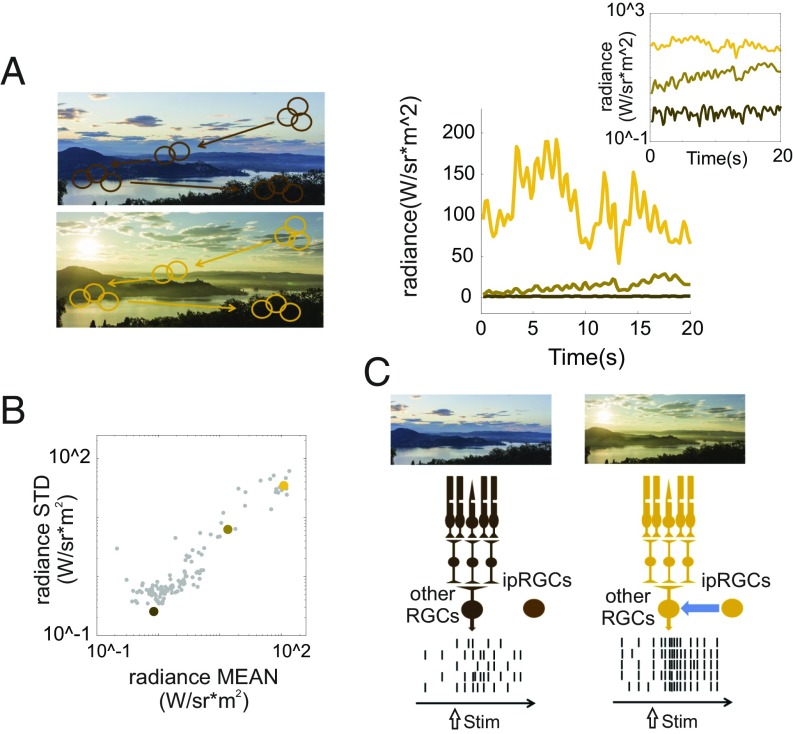
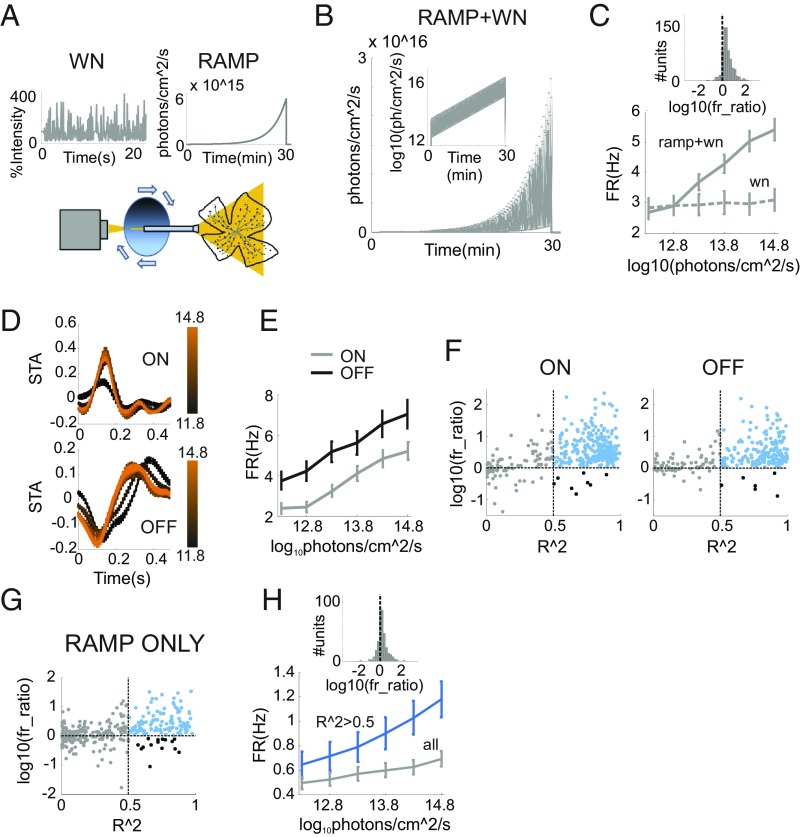
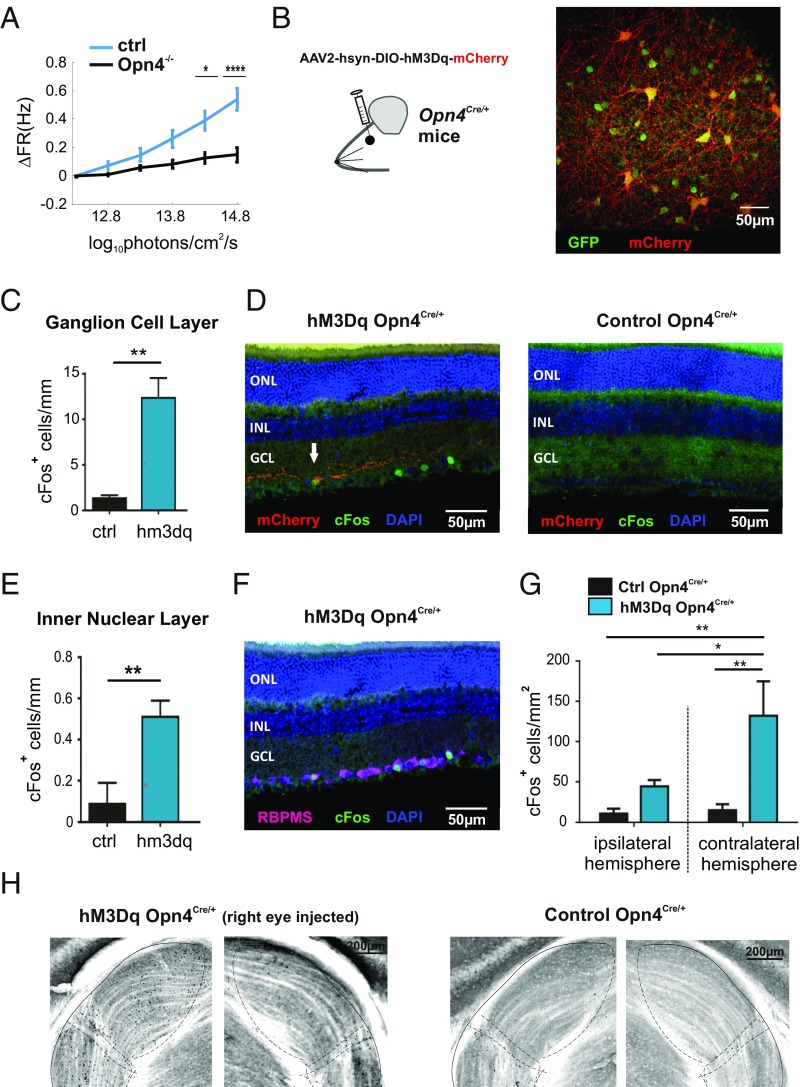
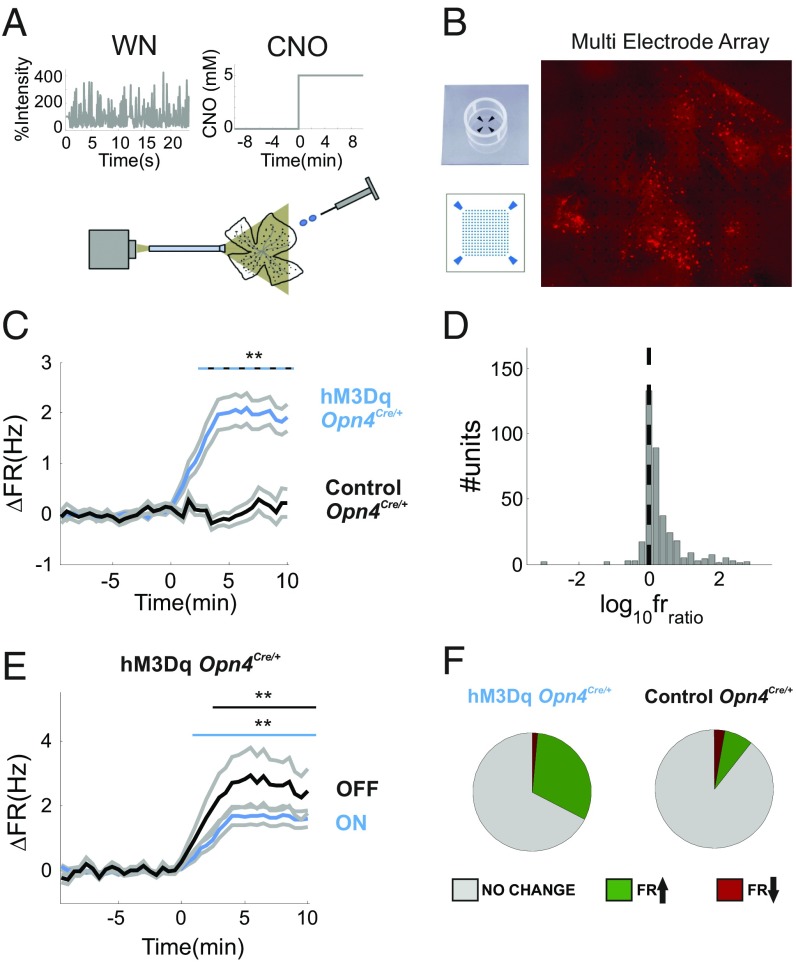
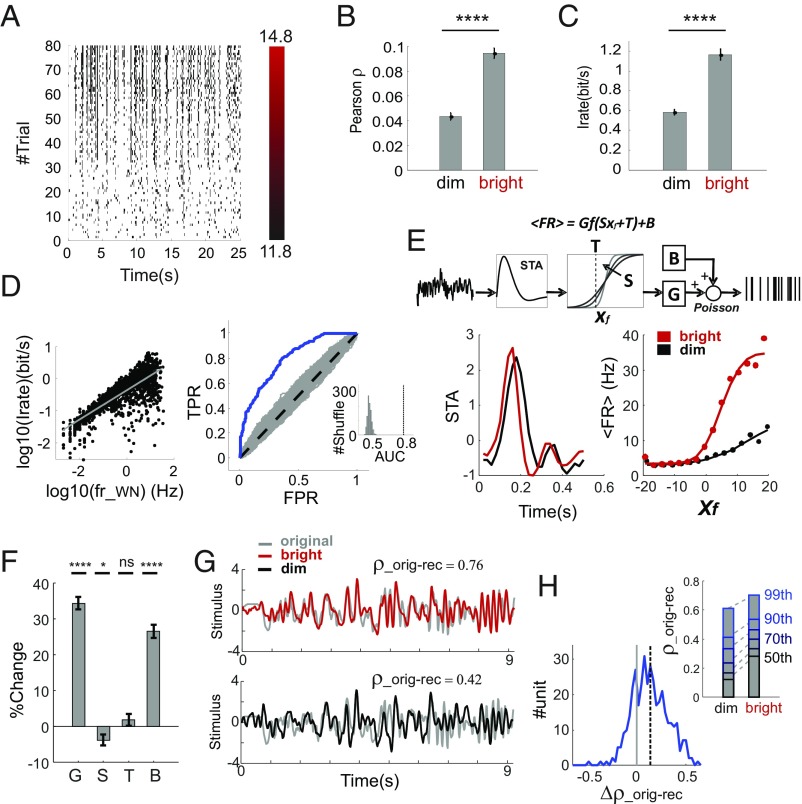
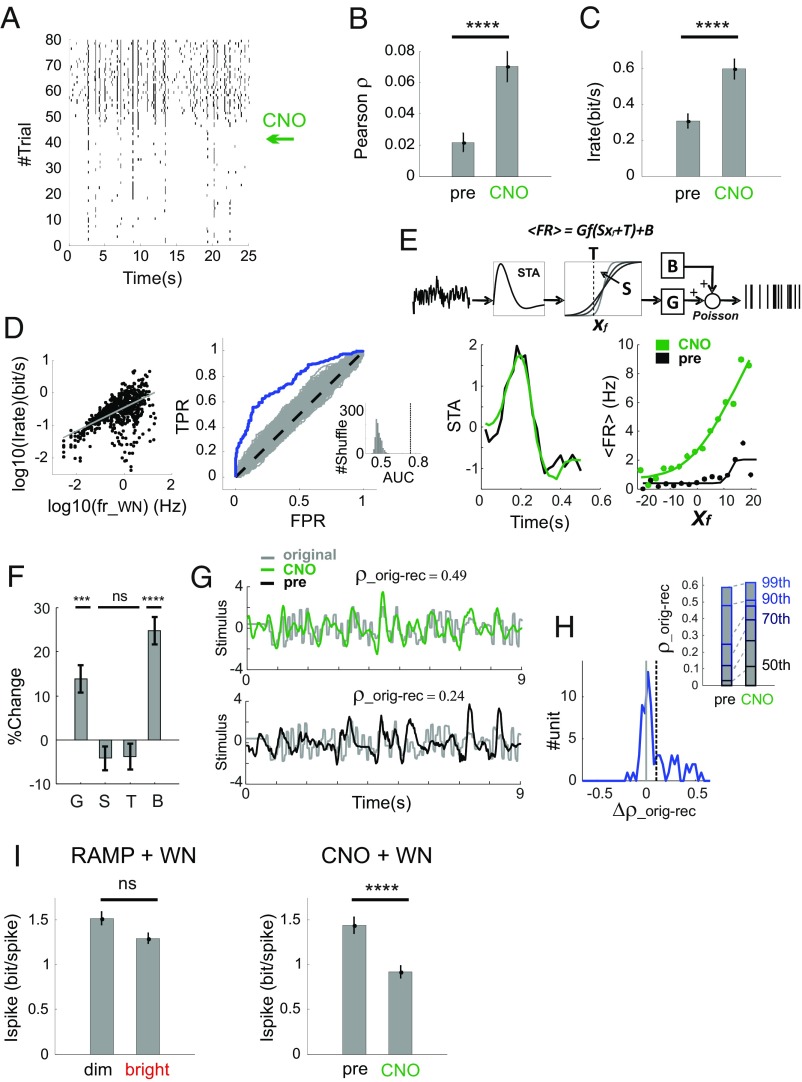
Information transfer in the brain relies upon energetically expensive spiking activity of neurons. Rates of information flow should therefore be carefully optimized, but mechanisms to control this parameter are poorly understood. We address this deficit in the visual system, where ambient light (irradiance) is predictive of the amount of information reaching the eye and ask whether a neural measure of irradiance can therefore be used to proactively control information flow along the optic nerve. We first show that firing rates for the retina's output neurons [retinal ganglion cells (RGCs)] scale with irradiance and are positively correlated with rates of information and the gain of visual responses. Irradiance modulates firing in the absence of any other visual signal confirming that this is a genuine response to changing ambient light. Irradiance-driven changes in firing are observed across the population of RGCs (including in both ON and OFF units) but are disrupted in mice lacking melanopsin [the photopigment of irradiance-coding intrinsically photosensitive RGCs (ipRGCs)] and can be induced under steady light exposure by chemogenetic activation of ipRGCs. Artificially elevating firing by chemogenetic excitation of ipRGCs is sufficient to increase information flow by increasing the gain of visual responses, indicating that enhanced firing is a cause of increased information transfer at higher irradiance. Our results establish a retinal circuitry driving changes in RGC firing as an active response to alterations in ambient light to adjust the amount of visual information transmitted to the brain.
Keywords: information; ipRGC; melanopsin; neural coding; retina.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
