

Liquid Nuclear Condensates Mechanically Sense and Restructure the Genome
- PMID: 30500535
- PMCID: PMC6724728
- DOI: 10.1016/j.cell.2018.10.057
Liquid Nuclear Condensates Mechanically Sense and Restructure the Genome
Erratum in
-
Liquid Nuclear Condensates Mechanically Sense and Restructure the Genome.Cell. 2019 Mar 7;176(6):1518. doi: 10.1016/j.cell.2019.02.025. Cell. 2019. PMID: 30849377 No abstract available.
Abstract
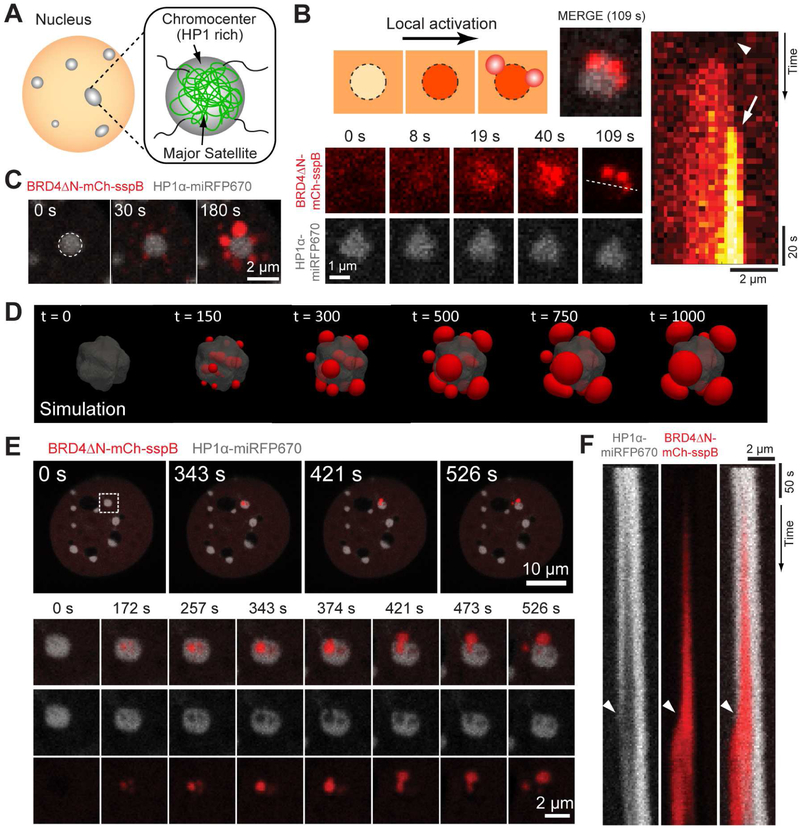
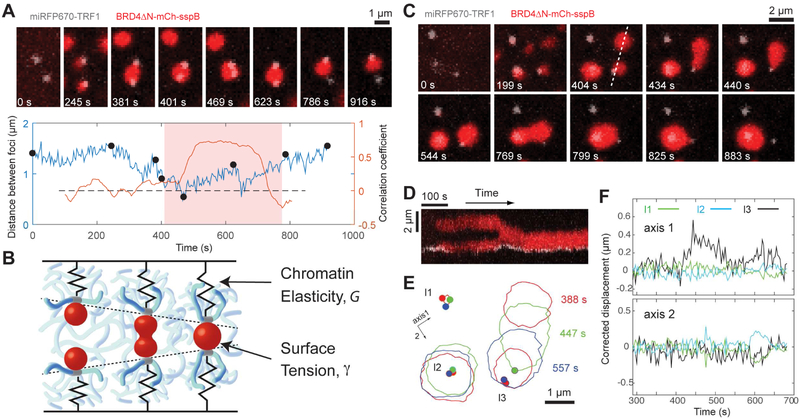
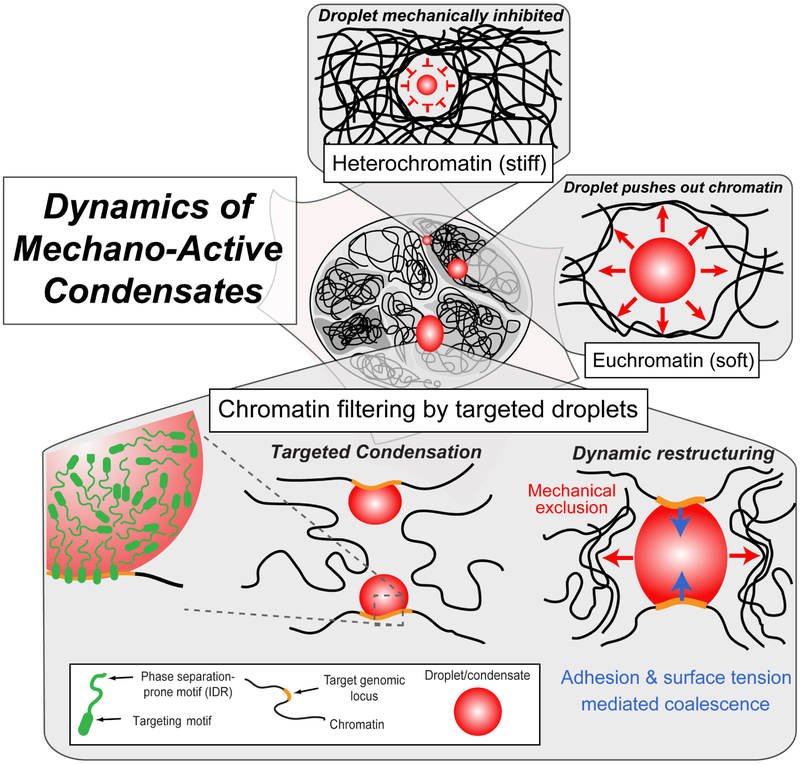
Phase transitions involving biomolecular liquids are a fundamental mechanism underlying intracellular organization. In the cell nucleus, liquid-liquid phase separation of intrinsically disordered proteins (IDPs) is implicated in assembly of the nucleolus, as well as transcriptional clusters, and other nuclear bodies. However, it remains unclear whether and how physical forces associated with nucleation, growth, and wetting of liquid condensates can directly restructure chromatin. Here, we use CasDrop, a novel CRISPR-Cas9-based optogenetic technology, to show that various IDPs phase separate into liquid condensates that mechanically exclude chromatin as they grow and preferentially form in low-density, largely euchromatic regions. A minimal physical model explains how this stiffness sensitivity arises from lower mechanical energy associated with deforming softer genomic regions. Targeted genomic loci can nonetheless be mechanically pulled together through surface tension-driven coalescence. Nuclear condensates may thus function as mechano-active chromatin filters, physically pulling in targeted genomic loci while pushing out non-targeted regions of the neighboring genome. VIDEO ABSTRACT.
Keywords: chromatin; condensates; gene regulation; mechanobiology; nuclear mechanics; nuclear organization; optogenetics; phase immiscibility; phase separation.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
A patent application describing the CasDrop technology is currently pending.
Figures
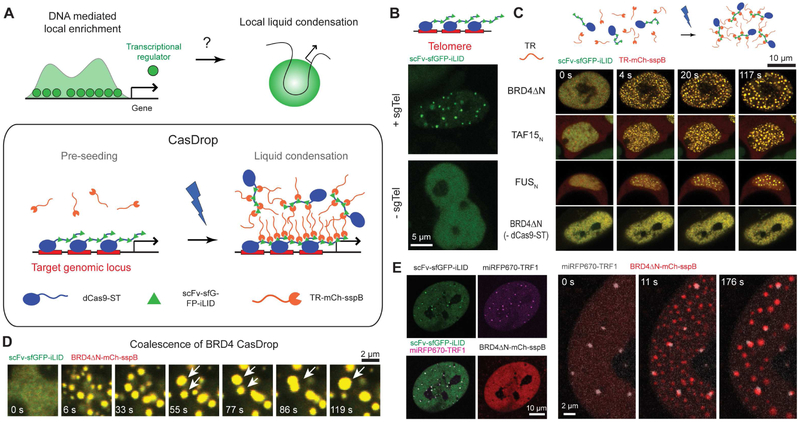
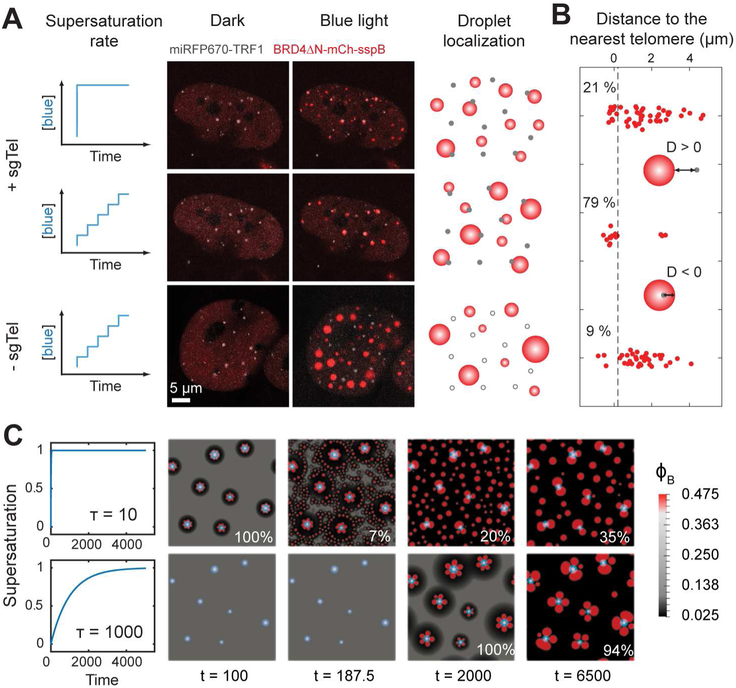
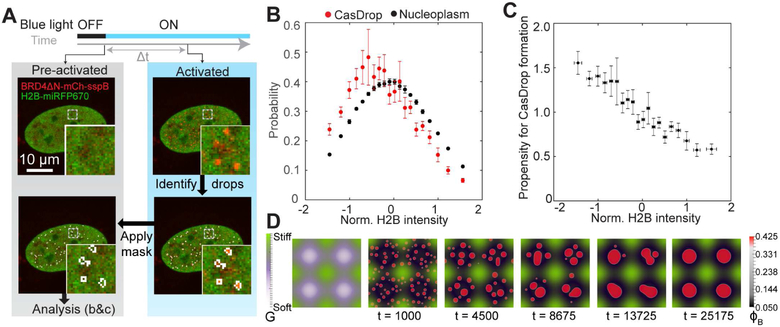
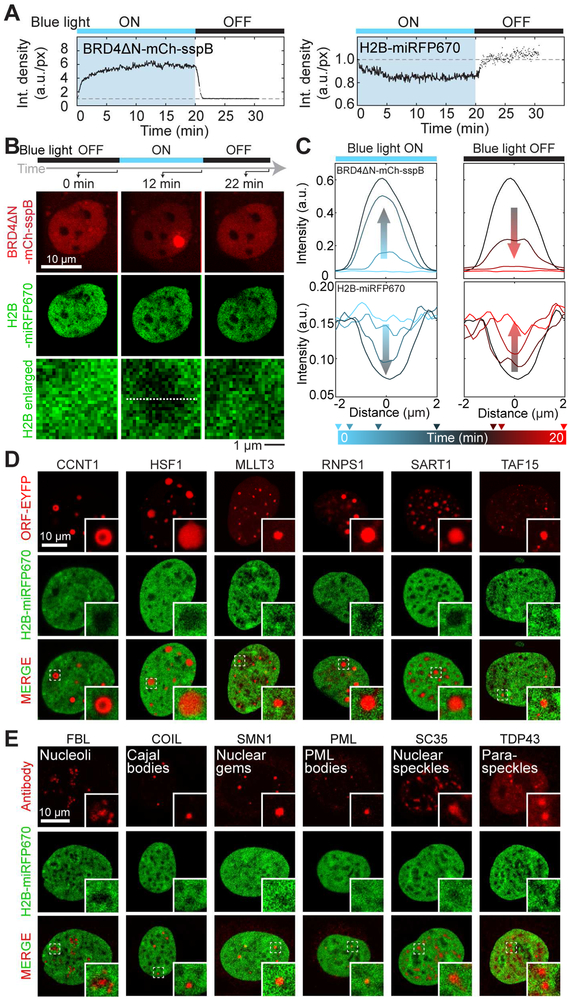
Comment in
-
Mechanobiology of Protein Droplets: Force Arises from Disorder.Cell. 2018 Nov 29;175(6):1457-1459. doi: 10.1016/j.cell.2018.11.020. Cell. 2018. PMID: 30500530
References
-
- Bracha D, Walls M, Wei M-T, Zhu L, Toettcher JE, and Brangwynne C (2018). Mapping local and global liquid-liquid phase behavior in living cells using light activated multivalent seeds. bioRxiv under review.
-
- Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Julicher F, and Hyman AA (2009). Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 324, 1729–1732. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
