

Role of the Mre11 Complex in Preserving Genome Integrity
- PMID: 30501098
- PMCID: PMC6315862
- DOI: 10.3390/genes9120589
Role of the Mre11 Complex in Preserving Genome Integrity
Abstract
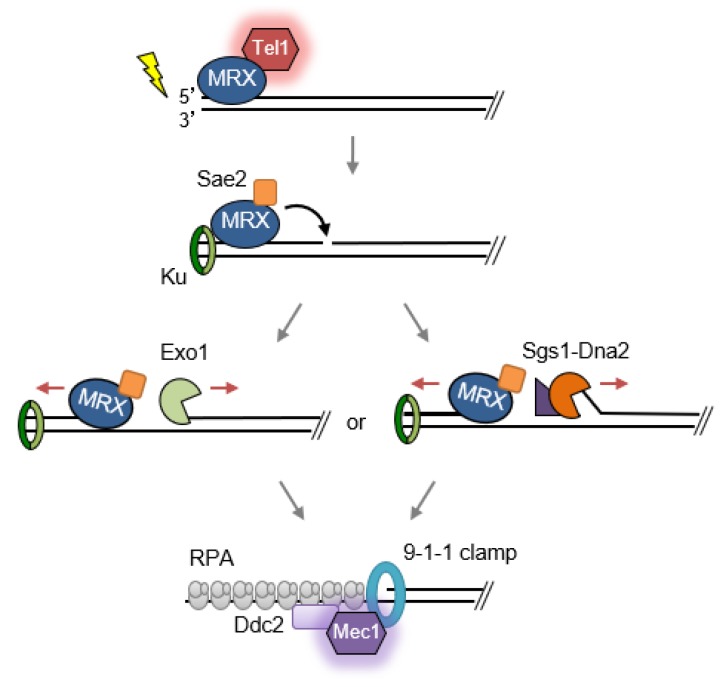
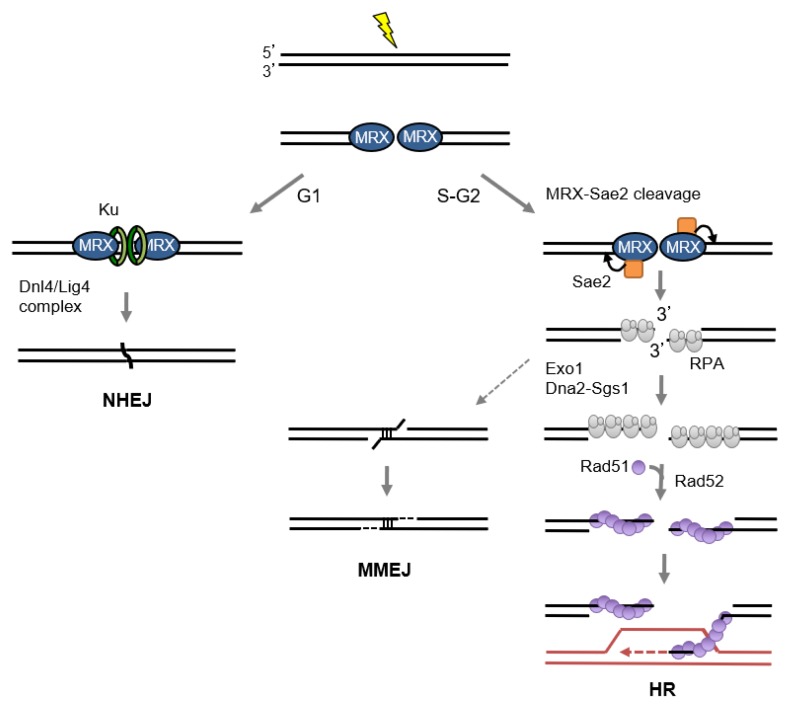
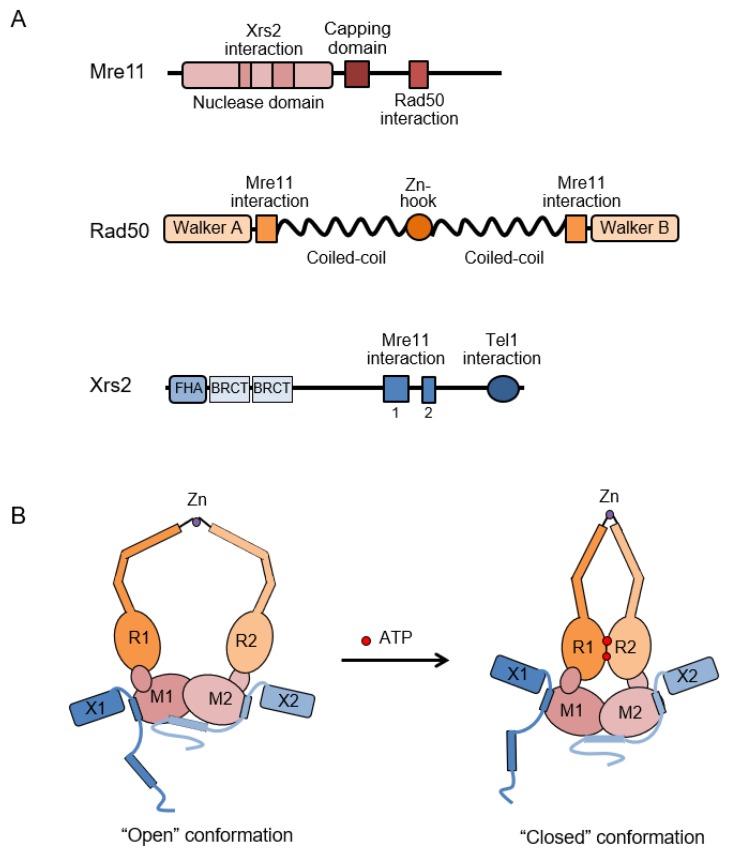
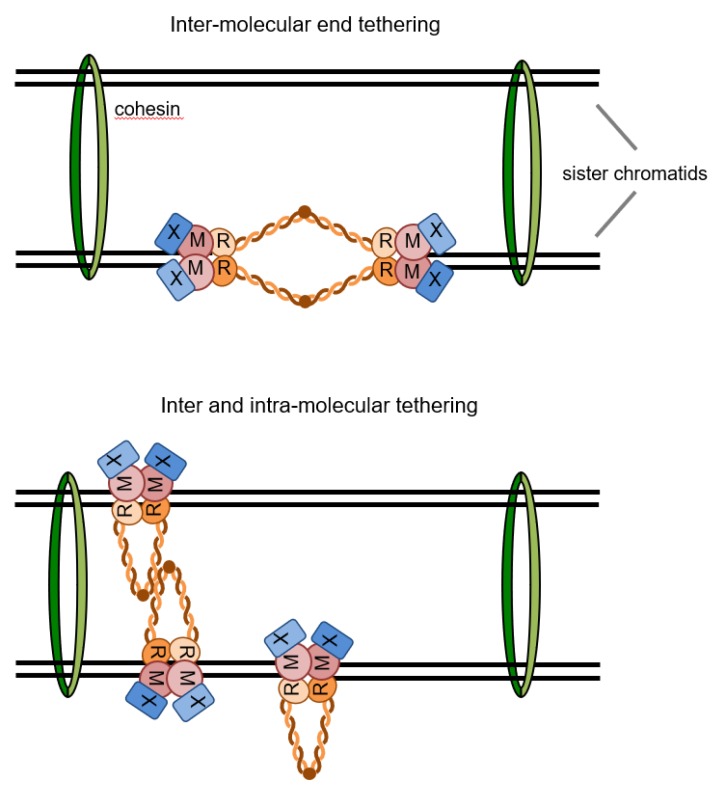
DNA double-strand breaks (DSBs) are hazardous lesions that threaten genome integrity and cell survival. The DNA damage response (DDR) safeguards the genome by sensing DSBs, halting cell cycle progression and promoting repair through either non-homologous end joining (NHEJ) or homologous recombination (HR). The Mre11-Rad50-Xrs2/Nbs1 (MRX/N) complex is central to the DDR through its structural, enzymatic, and signaling roles. The complex tethers DNA ends, activates the Tel1/ATM kinase, resolves protein-bound or hairpin-capped DNA ends, and maintains telomere homeostasis. In addition to its role at DSBs, MRX/N associates with unperturbed replication forks, as well as stalled replication forks, to ensure complete DNA synthesis and to prevent chromosome rearrangements. Here, we summarize the significant progress made in characterizing the MRX/N complex and its various activities in chromosome metabolism.
Keywords: DNA damage checkpoint; DNA repair; DSB; MRX/N; Mre11; Rad50; Sae2/Ctp1/CtIP; Tel1/ATM; Xrs2/Nbs1; homologous recombination.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Carney J.P., Maser R.S., Olivares H., Davis E.M., Le Beau M., Yates J.R., 3rd, Hays L., Morgan W.F., Petrini J.H. The hMre11/hRad50 protein complex and Nijmegen breakage syndrome: Linkage of double-strand break repair to the cellular DNA damage response. Cell. 1998;93:477–486. doi: 10.1016/S0092-8674(00)81175-7. - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
