

Cooperative Induction of Ordered Peptide and Fatty Acid Aggregates
- PMID: 30503535
- PMCID: PMC6302236
- DOI: 10.1016/j.bpj.2018.10.031
Cooperative Induction of Ordered Peptide and Fatty Acid Aggregates
Abstract
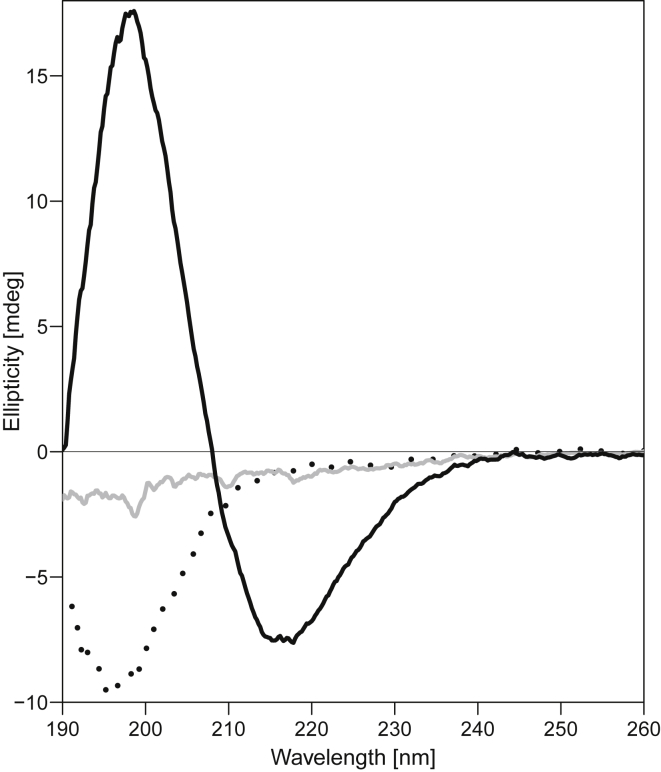
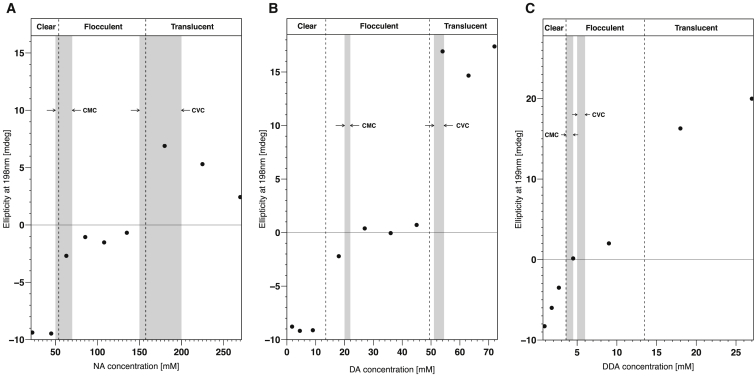
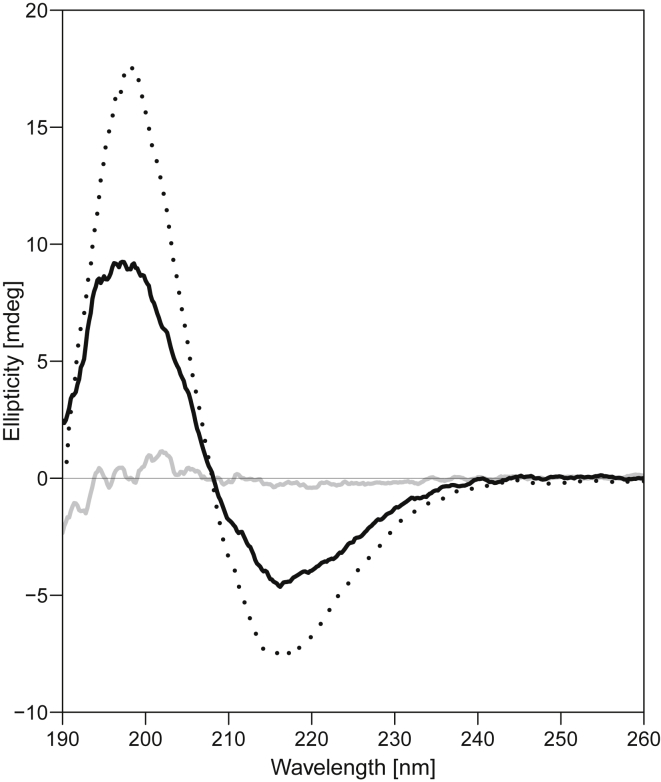
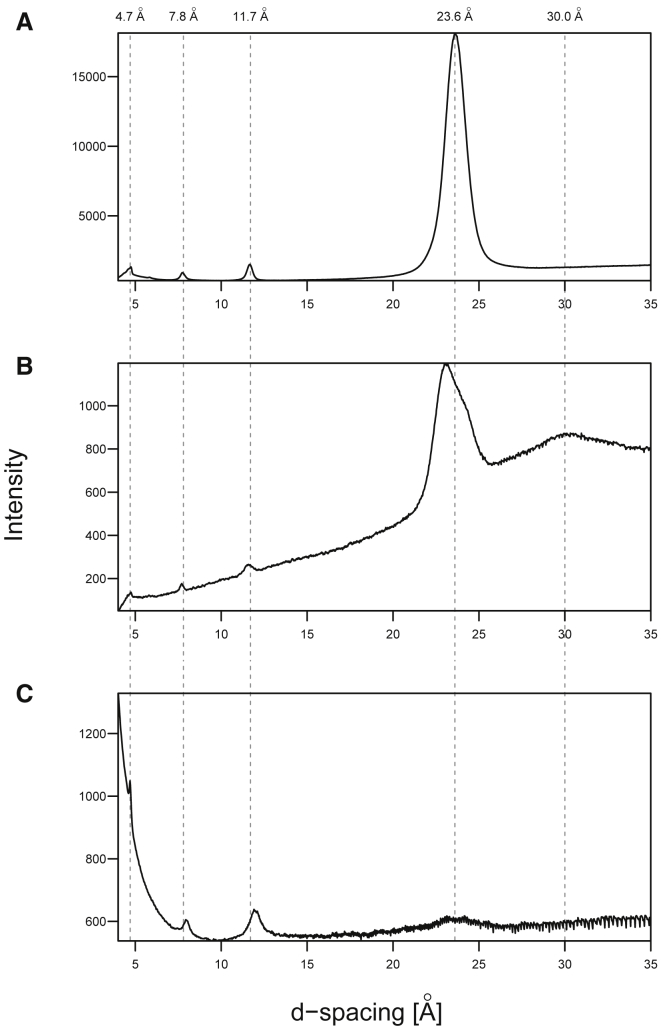
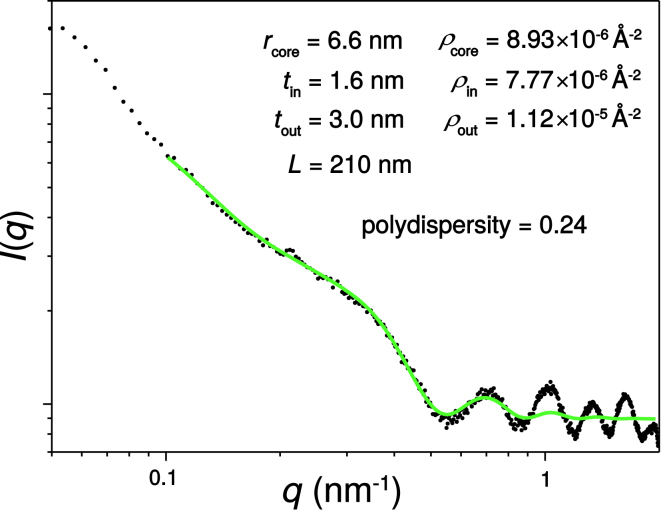
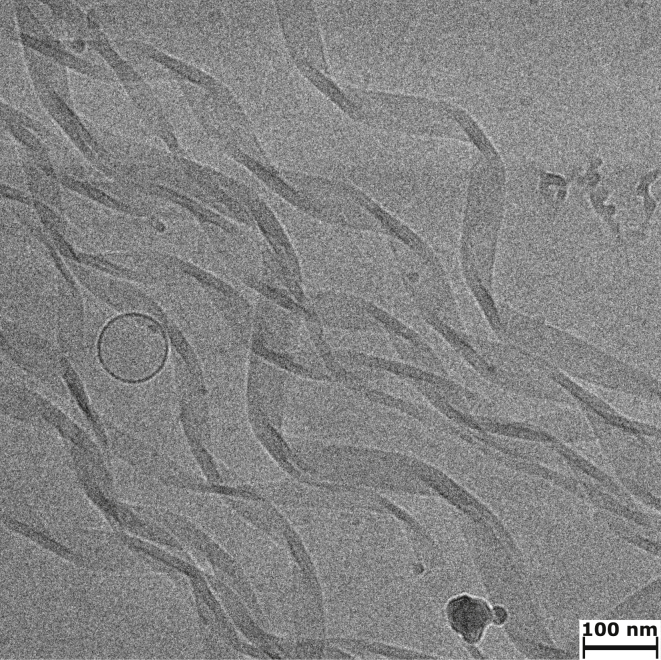
Interactions between biological membranes and disease-associated amyloids are well documented, and their prevalence suggests that an inherent affinity exists between these molecular assemblies. Our interest in the molecular origins of life have led us to investigate the nature of such interactions in the context of their molecular predecessors (i.e., vesicle-forming amphiphiles and small peptides). Under certain conditions, amyloidogenic peptides or fatty acids are each able to form ordered structures on their own; however, we report here on their cooperative assembly into novel, to our knowledge, highly ordered structures. We first examined an amyloidogenic eight-residue peptide, which forms amyloids at pH 11, yet because of its positive electrostatic character remains soluble at a neutral pH. In mixtures with simple fatty acids, this peptide is also able to form novel, to our knowledge, coaggregates at a neutral pH whose structures are sensitive to both the fatty acid concentration and identity. Below the critical vesicle concentration, the mixtures of fatty acid and peptide yield a flocculent precipitate with an underlying β-structure. Above the critical vesicle concentration, the mixtures yield a translucent precipitate that consists of tube-like structures. Small-angle x-ray scattering and fiber diffraction data were used to model their structures as hollow-core two-shell cylinders in which the inner shell is a bilayer of fatty acid and the outer shell alternates between amyloid and bilayers of fatty acid. The further analysis of decanoic acid with a panel of 13 other basic amyloidogenic peptides confirmed the general nature of the observed interactions. The cooperativity within this heterogeneous system is attributed to the structurally repetitive natures of the fatty acid bilayer and the cross-β-sheet motif, providing compatible scaffolds for attractive electrostatic interactions. We show these interactions to be mutually beneficial, expanding the phase space of both peptides and fatty acids while providing a simple yet robust physical connection between two distinct entities relevant for life.
Copyright © 2018 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures

References
-
- Orgel L.E. Evolution of the genetic apparatus. J. Mol. Biol. 1968;38:381–393. - PubMed
-
- Segré D., Ben-Eli D., Lancet D. The lipid world. Orig. Life Evol. Biosph. 2001;31:119–145. - PubMed
-
- Gilbert W. Origin of life: the RNA world. Nature. 1986;319:618.
-
- Oba T., Fukushima J., Ikehara K. Catalytic activities of [GADV]-peptides. Formation and establishment of [GADV]-protein world for the emergence of life. Orig. Life Evol. Biosph. 2005;35:447–460. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
