

Widespread Backtracking by RNA Pol II Is a Major Effector of Gene Activation, 5' Pause Release, Termination, and Transcription Elongation Rate
- PMID: 30503775
- PMCID: PMC6320282
- DOI: 10.1016/j.molcel.2018.10.031
Widespread Backtracking by RNA Pol II Is a Major Effector of Gene Activation, 5' Pause Release, Termination, and Transcription Elongation Rate
Abstract
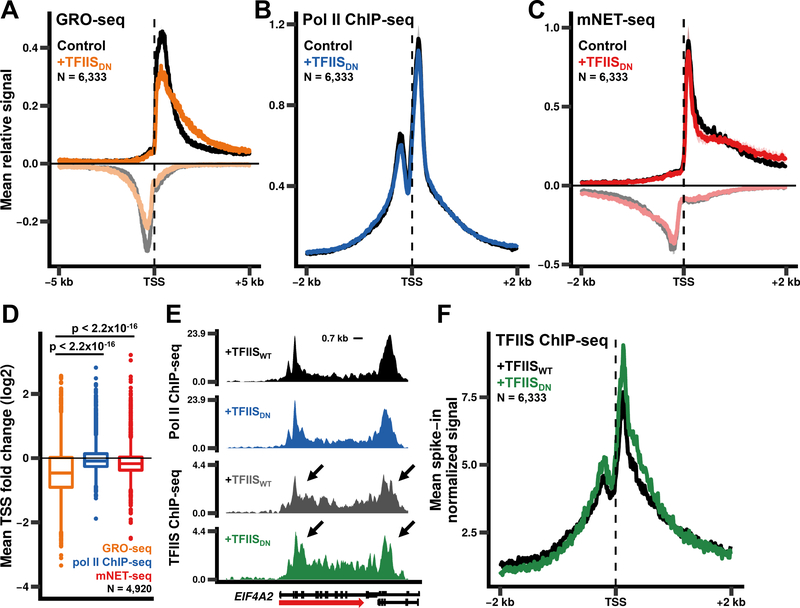
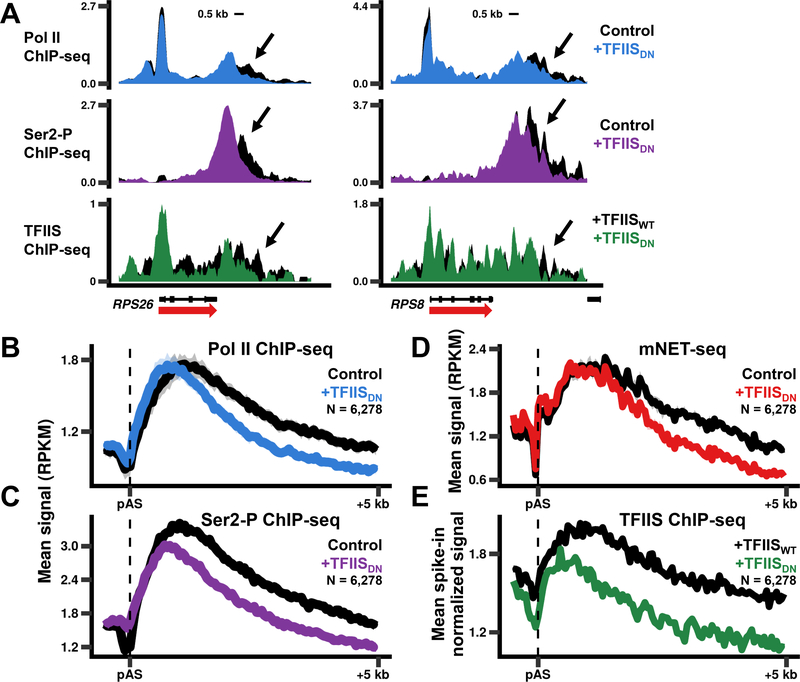
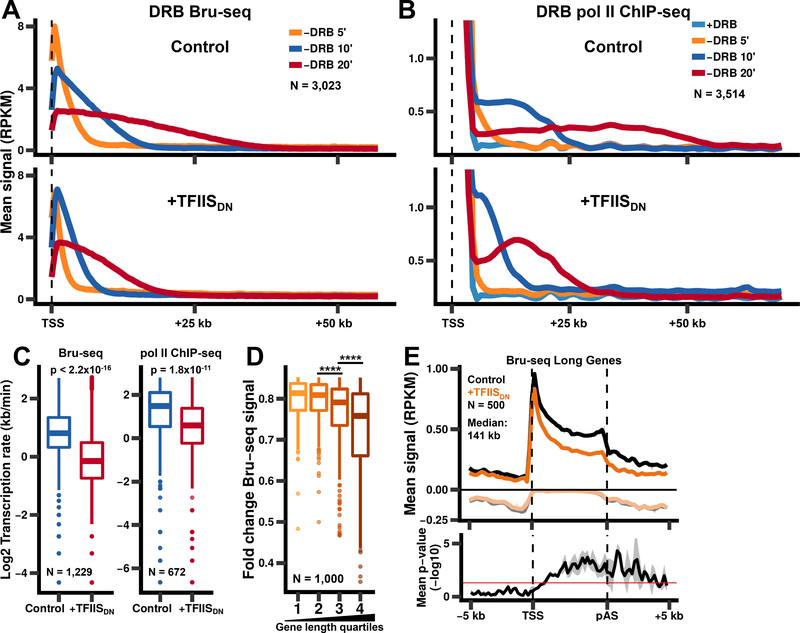
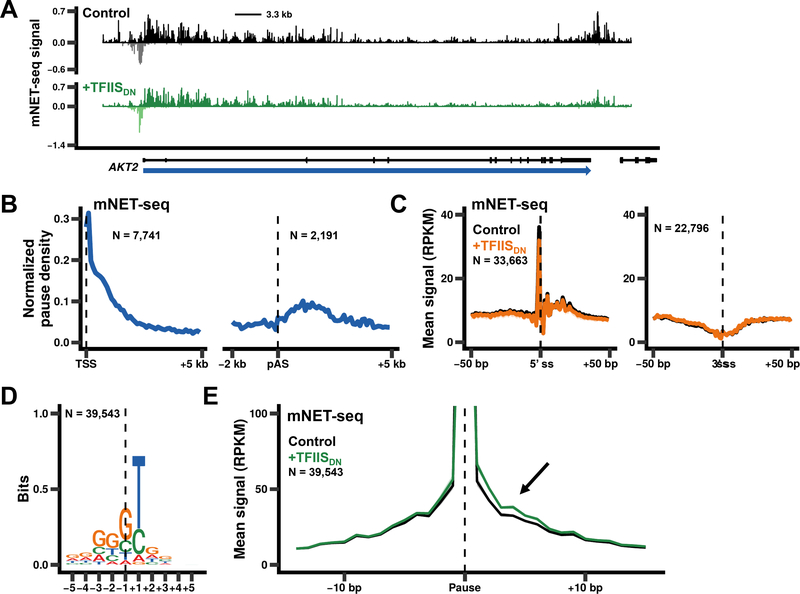
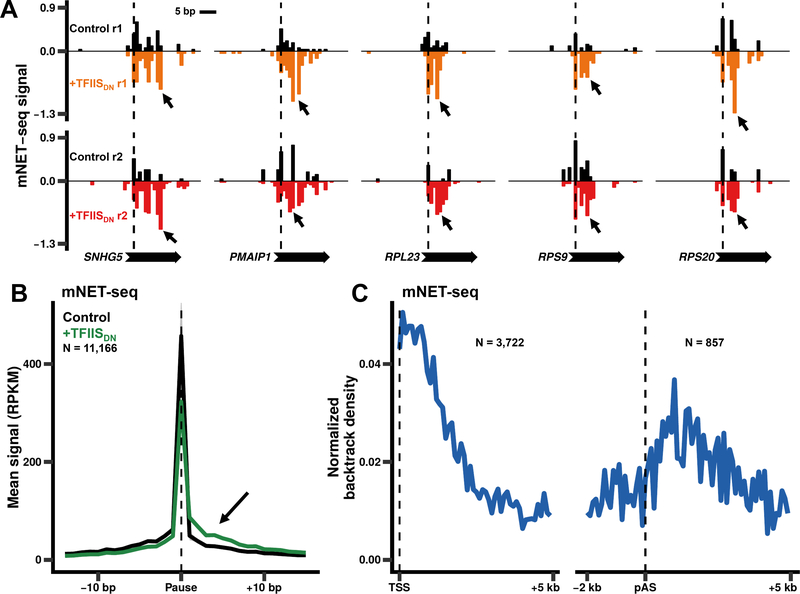
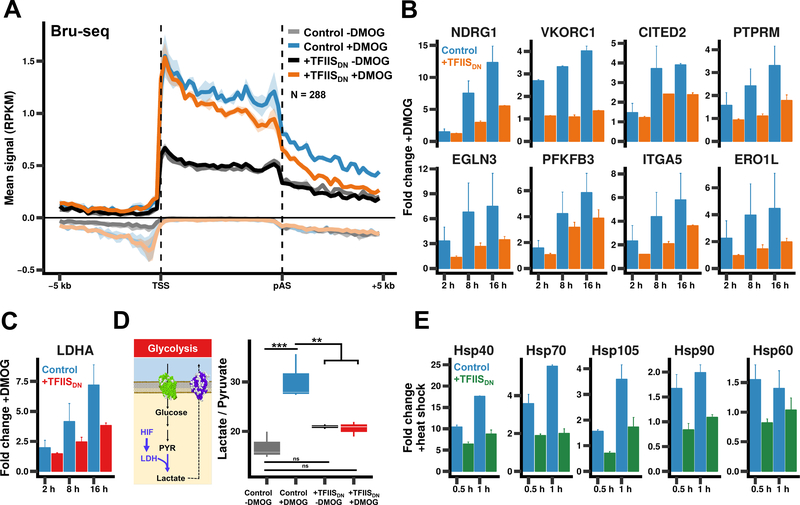
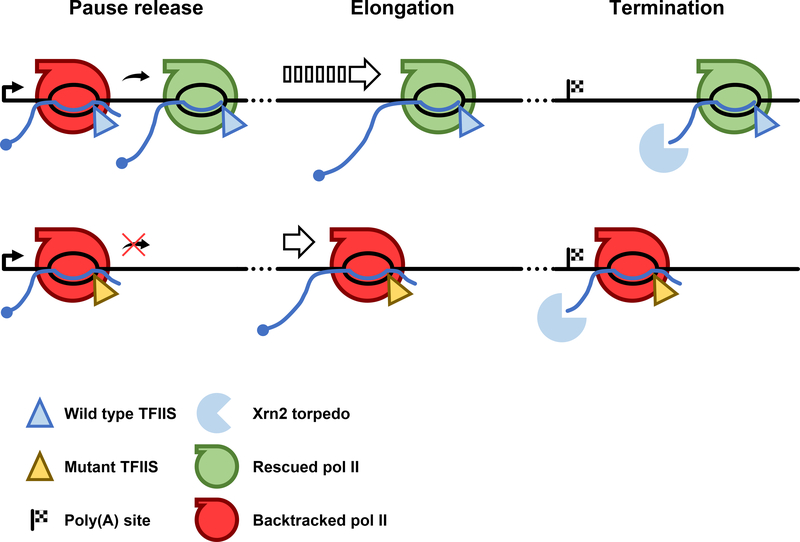
In addition to phosphodiester bond formation, RNA polymerase II has an RNA endonuclease activity, stimulated by TFIIS, which rescues complexes that have arrested and backtracked. How TFIIS affects transcription under normal conditions is poorly understood. We identified backtracking sites in human cells using a dominant-negative TFIIS (TFIISDN) that inhibits RNA cleavage and stabilizes backtracked complexes. Backtracking is most frequent within 2 kb of start sites, consistent with slow elongation early in transcription, and in 3' flanking regions where termination is enhanced by TFIISDN, suggesting that backtracked pol II is a favorable substrate for termination. Rescue from backtracking by RNA cleavage also promotes escape from 5' pause sites, prevents premature termination of long transcripts, and enhances activation of stress-inducible genes. TFIISDN slowed elongation rates genome-wide by half, suggesting that rescue of backtracked pol II by TFIIS is a major stimulus of elongation under normal conditions.
Keywords: RNA polymerase pausing; TFIIS; hypoxia response; transcription elongation; transcription termination; transcriptional backtracking.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests
Authors declare no competing interests.
Figures
Similar articles
-
Transcription factors TFIIF and TFIIS promote transcript elongation by RNA polymerase II by synergistic and independent mechanisms.Proc Natl Acad Sci U S A. 2014 May 6;111(18):6642-7. doi: 10.1073/pnas.1405181111. Epub 2014 Apr 14. Proc Natl Acad Sci U S A. 2014. PMID: 24733897 Free PMC article.
-
Nature of the nucleosomal barrier to RNA polymerase II.Mol Cell. 2005 Apr 1;18(1):97-108. doi: 10.1016/j.molcel.2005.02.027. Mol Cell. 2005. PMID: 15808512
-
Causes and consequences of RNA polymerase II stalling during transcript elongation.Nat Rev Mol Cell Biol. 2021 Jan;22(1):3-21. doi: 10.1038/s41580-020-00308-8. Epub 2020 Nov 18. Nat Rev Mol Cell Biol. 2021. PMID: 33208928 Review.
-
Elongation Factor TFIIS Prevents Transcription Stress and R-Loop Accumulation to Maintain Genome Stability.Mol Cell. 2019 Oct 3;76(1):57-69.e9. doi: 10.1016/j.molcel.2019.07.037. Epub 2019 Sep 10. Mol Cell. 2019. PMID: 31519522 Free PMC article.
-
Pause Patrol: Negative Elongation Factor's Role in Promoter-Proximal Pausing and Beyond.J Mol Biol. 2025 Jan 1;437(1):168779. doi: 10.1016/j.jmb.2024.168779. Epub 2024 Sep 4. J Mol Biol. 2025. PMID: 39241983 Review.
Cited by
-
Control of RNA Pol II Speed by PNUTS-PP1 and Spt5 Dephosphorylation Facilitates Termination by a "Sitting Duck Torpedo" Mechanism.Mol Cell. 2019 Dec 19;76(6):896-908.e4. doi: 10.1016/j.molcel.2019.09.031. Epub 2019 Oct 30. Mol Cell. 2019. PMID: 31677974 Free PMC article.
-
Organismal benefits of transcription speed control at gene boundaries.EMBO Rep. 2020 Apr 3;21(4):e49315. doi: 10.15252/embr.201949315. Epub 2020 Feb 27. EMBO Rep. 2020. PMID: 32103605 Free PMC article.
-
Transcription stress at telomeres leads to cytosolic DNA release and paracrine senescence.Nat Commun. 2024 May 14;15(1):4061. doi: 10.1038/s41467-024-48443-6. Nat Commun. 2024. PMID: 38744897 Free PMC article.
-
Integrated genome and transcriptome analyses reveal the mechanism of genome instability in ataxia with oculomotor apraxia 2.Proc Natl Acad Sci U S A. 2022 Jan 25;119(4):e2114314119. doi: 10.1073/pnas.2114314119. Proc Natl Acad Sci U S A. 2022. PMID: 35042798 Free PMC article.
-
RNA Polymerase II Activity Control of Gene Expression and Involvement in Disease.J Mol Biol. 2025 Jan 1;437(1):168770. doi: 10.1016/j.jmb.2024.168770. Epub 2024 Aug 28. J Mol Biol. 2025. PMID: 39214283 Free PMC article. Review.
References
-
- Adelman K, Marr MT, Werner J, Saunders A, Ni Z, Andrulis ED, and Lis JT (2005). Efficient release from promoter-proximal stall sites requires transcript cleavage factor TFIIS. Mol Cell 17, 103112. - PubMed
-
- Bondarenko VA, Steele LM, Ujvari A, Gaykalova DA, Kulaeva OI, Polikanov YS, Luse DS, and Studitsky VM (2006). Nucleosomes can form a polar barrier to transcript elongation by RNA polymerase II. Mol Cell 24, 469–479. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
