

Engineering an Osmosensor by Pivotal Histidine Positioning within Disordered Helices
- PMID: 30503779
- PMCID: PMC6377431
- DOI: 10.1016/j.str.2018.10.012
Engineering an Osmosensor by Pivotal Histidine Positioning within Disordered Helices
Abstract
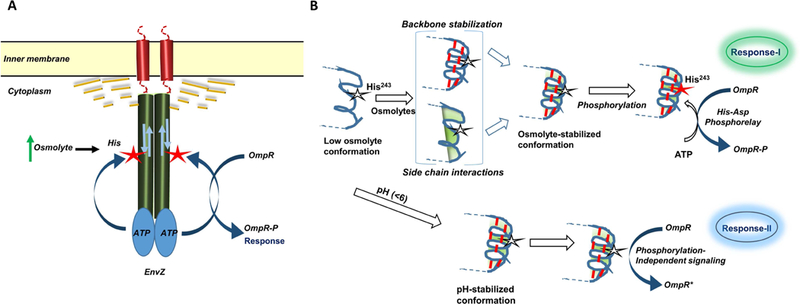
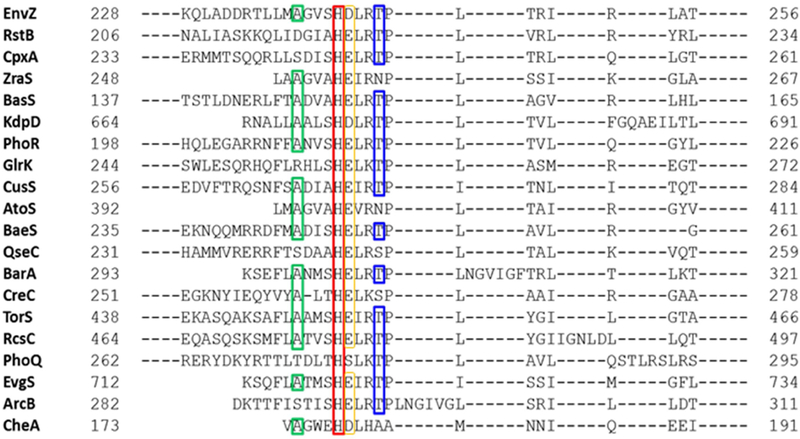
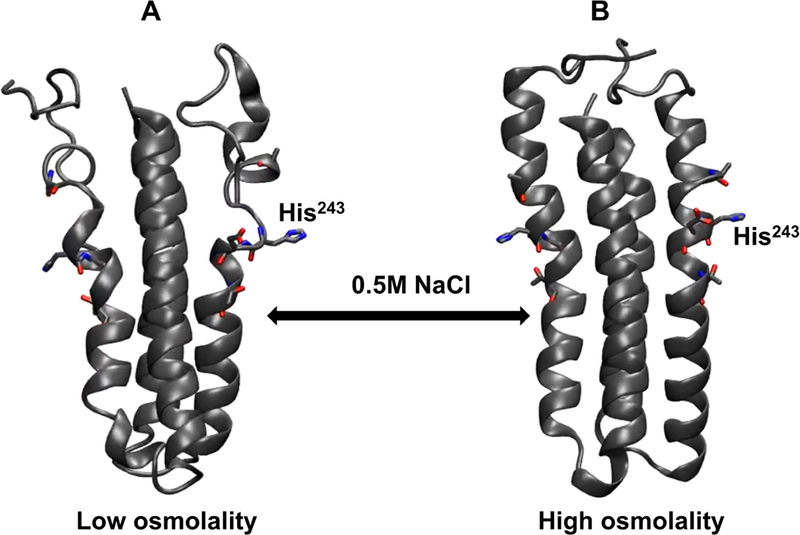
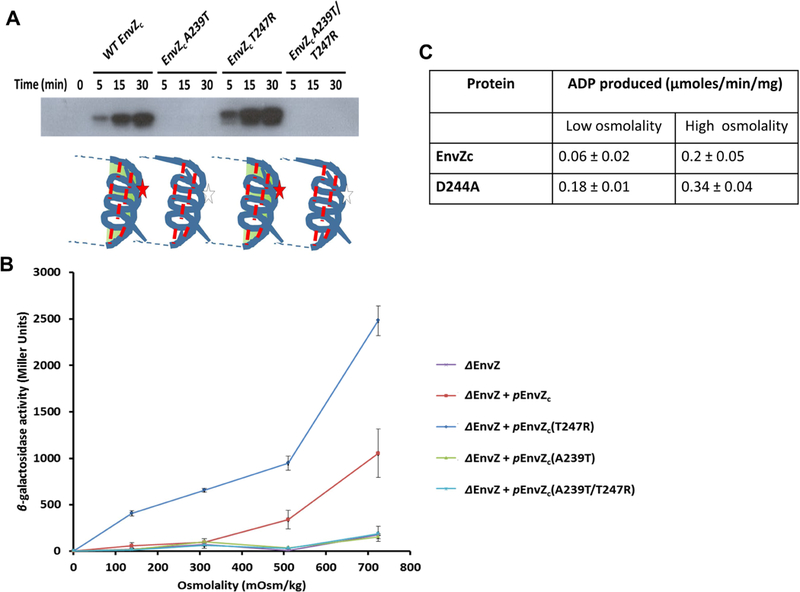
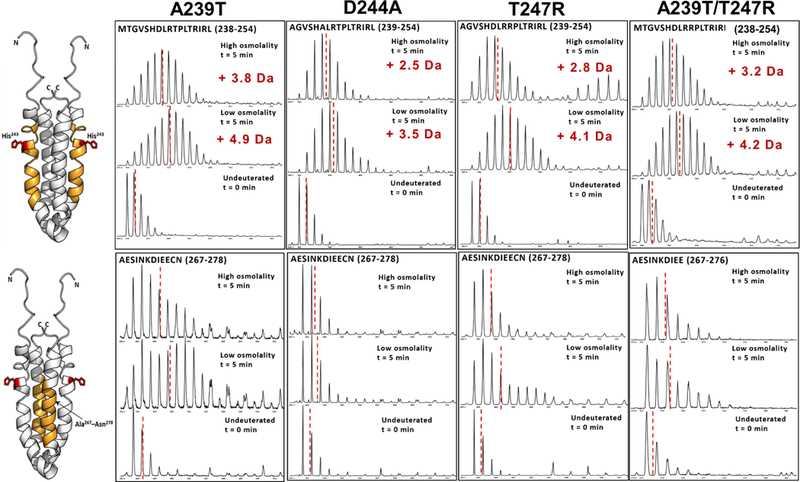
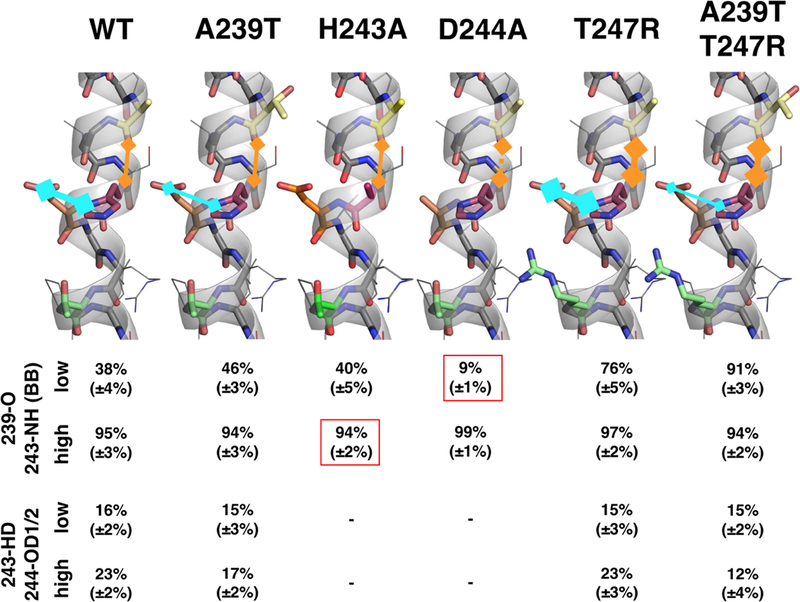
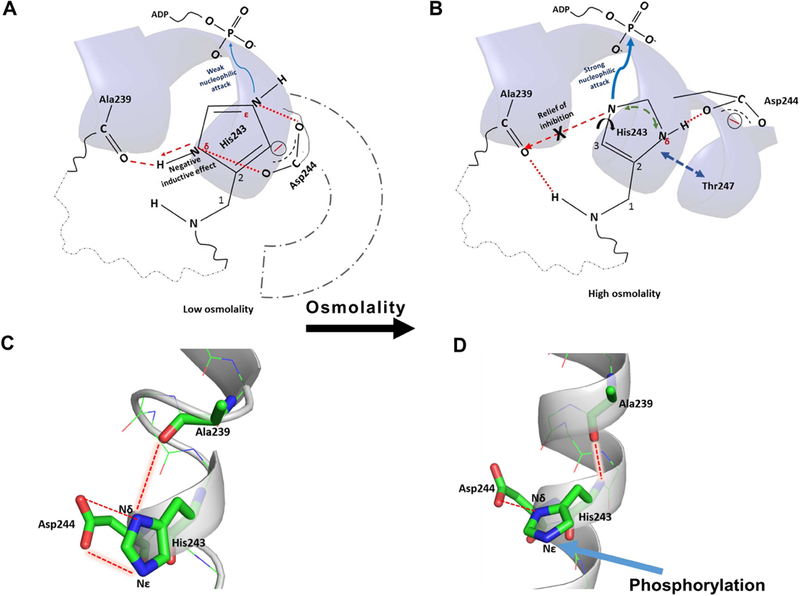
Histidine kinases (HKs) funnel diverse environmental stimuli into a single autophosphorylation event at a conserved histidine residue. The HK EnvZ is a global sensor of osmolality and cellular acid pH. In previous studies, we discovered that osmosensing in EnvZ was mediated through osmolyte-induced stabilization of the partially disordered helical backbone spanning the conserved histidine autophosphorylation site (His243). Here, we describe how backbone stabilization leads to changes in the microenvironment of His243, resulting in enhanced autophosphorylation through relief of inhibition and repositioning of critical side chains and imidazole rotamerization. The conserved His-Asp/Glu dyad within the partially structured helix is equally geared to respond to acid pH, an alternative environmental stimulus in bacteria. This high-resolution "double-clamp" switch model proposes that a His-Asp/Glu dyad functions as an integrative node for regulating autophosphorylation in HKs. Because the His-Asp/Glu dyad is highly conserved in HKs, this study provides a universal model for describing HK function.
Keywords: EnvZ; H bonding; His-Asp/Glu dyad; amide hydrogen-deuterium exchange mass spectrometry; helix stabilization; histidine rotamerization; osmosensing; protein dynamics; two-component signal transduction.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
DECLARATION OF INTEREST
The authors report no conflict of interest with the findings reported in this manuscript.
Figures
References
-
- Antommattei FM, and Weis RM (2006). 12 Reversible methylation of glutamate residues in the receptor proteins of bacterial sensory systems. Enzymes 24, 325–382. - PubMed
-
- Baron R, and McCammon JA (2013). Molecular recognition and ligand association. Annual Review of Physical Chemistry 64, 151–175. - PubMed
-
- Beier D, and Gross R (2006). Regulation of bacterial virulence by two-component systems. Curr Opin Microbiol 9, 143–152. - PubMed
-
- Berendsen HJC, van der Spoel D, and van Drunen R (1995). GROMACS: A message-passing parallel molecular dynamics implementation. Computer Physics Communications 91, 43–56.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
