

The interface between phosphatidylinositol transfer protein function and phosphoinositide signaling in higher eukaryotes
- PMID: 30504233
- PMCID: PMC6358302
- DOI: 10.1194/jlr.R089730
The interface between phosphatidylinositol transfer protein function and phosphoinositide signaling in higher eukaryotes
Abstract
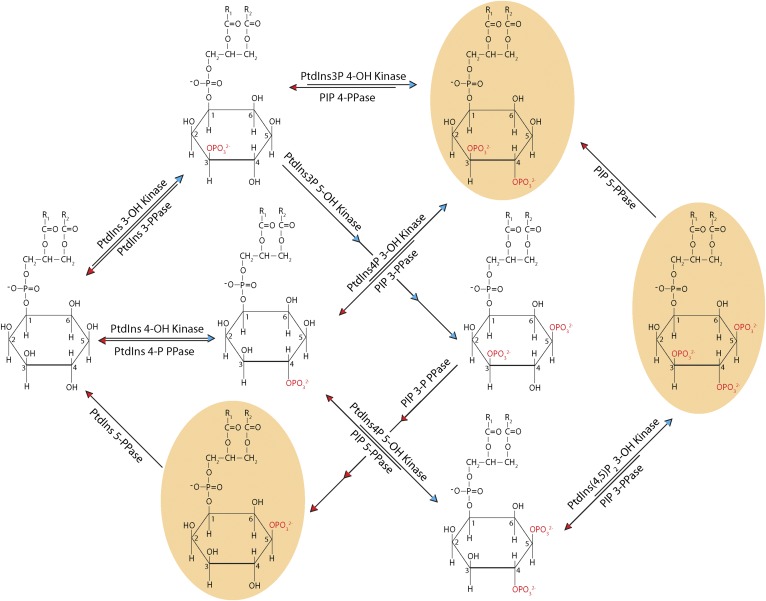
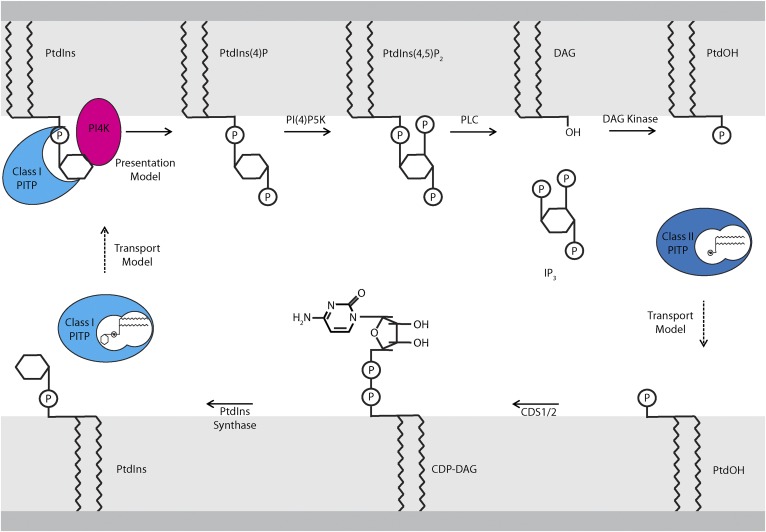
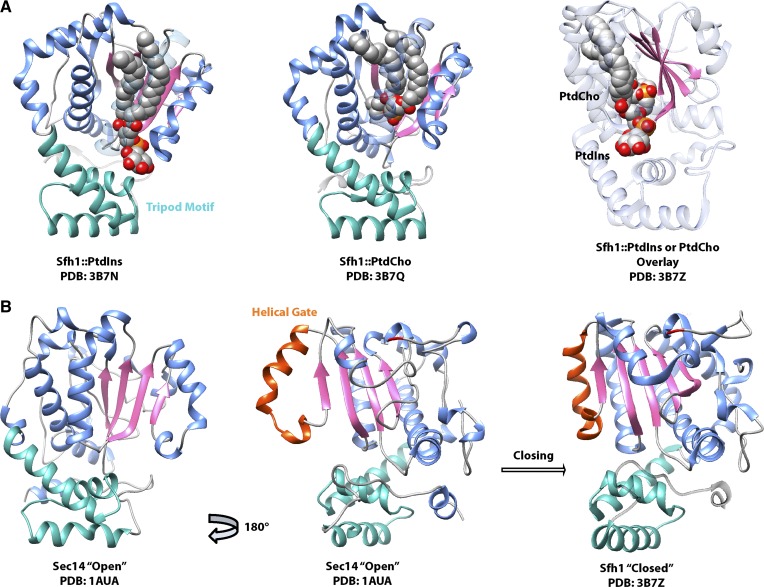
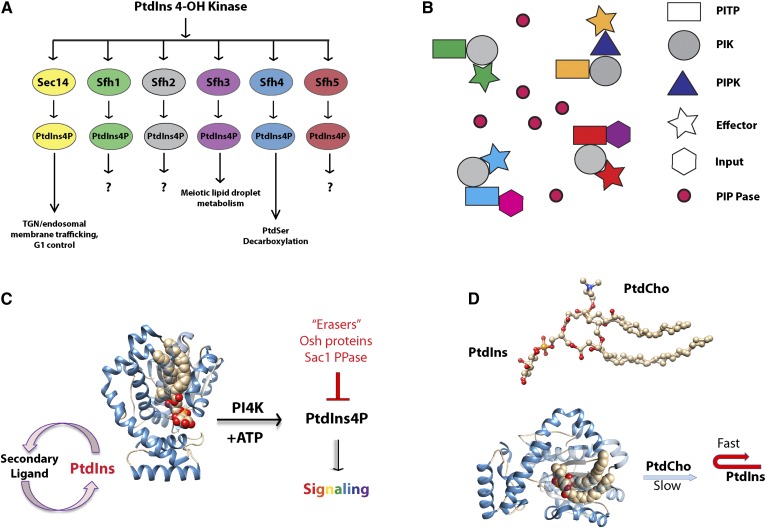
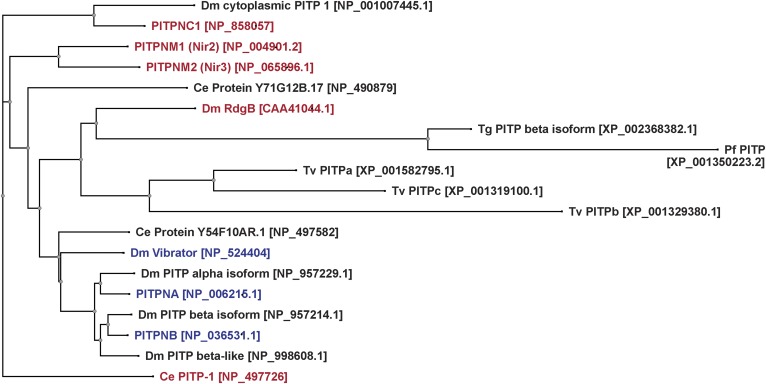
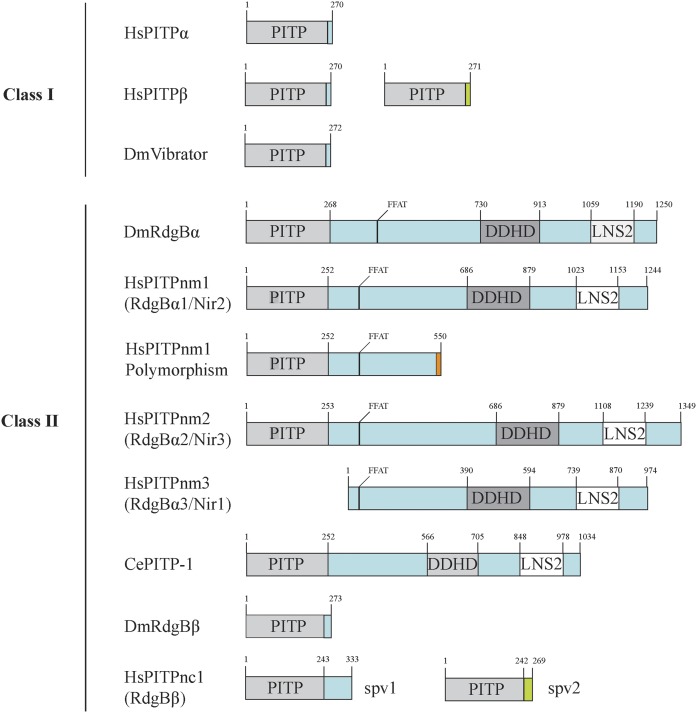
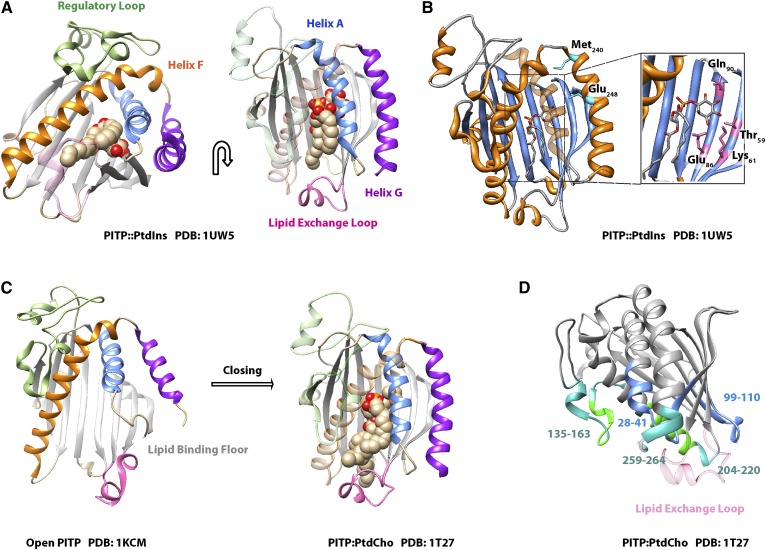
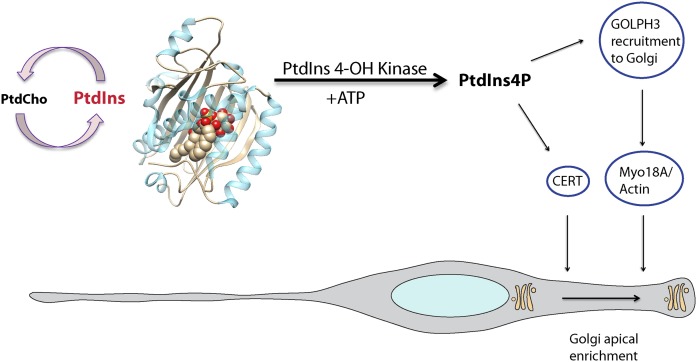
Phosphoinositides are key regulators of a large number of diverse cellular processes that include membrane trafficking, plasma membrane receptor signaling, cell proliferation, and transcription. How a small number of chemically distinct phosphoinositide signals are functionally amplified to exert specific control over such a diverse set of biological outcomes remains incompletely understood. To this end, a novel mechanism is now taking shape, and it involves phosphatidylinositol (PtdIns) transfer proteins (PITPs). The concept that PITPs exert instructive regulation of PtdIns 4-OH kinase activities and thereby channel phosphoinositide production to specific biological outcomes, identifies PITPs as central factors in the diversification of phosphoinositide signaling. There are two evolutionarily distinct families of PITPs: the Sec14-like and the StAR-related lipid transfer domain (START)-like families. Of these two families, the START-like PITPs are the least understood. Herein, we review recent insights into the biochemical, cellular, and physiological function of both PITP families with greater emphasis on the START-like PITPs, and we discuss the underlying mechanisms through which these proteins regulate phosphoinositide signaling and how these actions translate to human health and disease.
Keywords: cell signaling; diseases; lipid and membrane trafficking; lipid signaling; lipids • membranes.
Copyright © 2019 Grabon et al.
Figures
References
-
- Michell R. H. 2008. Inositol derivatives: evolution and functions. Nat. Rev. Mol. Cell Biol. 9: 151–161. - PubMed
-
- Michell R. H. 2013. Inositol lipids: from an archaeal origin to phosphatidylinositol 3,5-bisphosphate faults in human disease. FEBS J. 280: 6281–6294. - PubMed
-
- Behnia R., and Munro S.. 2005. Organelle identity and the signposts for membrane traffic. Nature. 438: 597–604. - PubMed
-
- Hokin M. R., and Hokin L. E.. 1954. Effects of acetylcholine on phospholipides in the pancreas. J. Biol. Chem. 209: 549–558. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
