

Systematically investigating the key features of the DNase deactivated Cpf1 for tunable transcription regulation in prokaryotic cells
- PMID: 30505961
- PMCID: PMC6251280
- DOI: 10.1016/j.synbio.2018.11.002
Systematically investigating the key features of the DNase deactivated Cpf1 for tunable transcription regulation in prokaryotic cells
Erratum in
-
Erratum regarding previously published articles.Synth Syst Biotechnol. 2020 Oct 12;5(4):328. doi: 10.1016/j.synbio.2020.10.003. eCollection 2020 Dec. Synth Syst Biotechnol. 2020. PMID: 33102826 Free PMC article.
Abstract
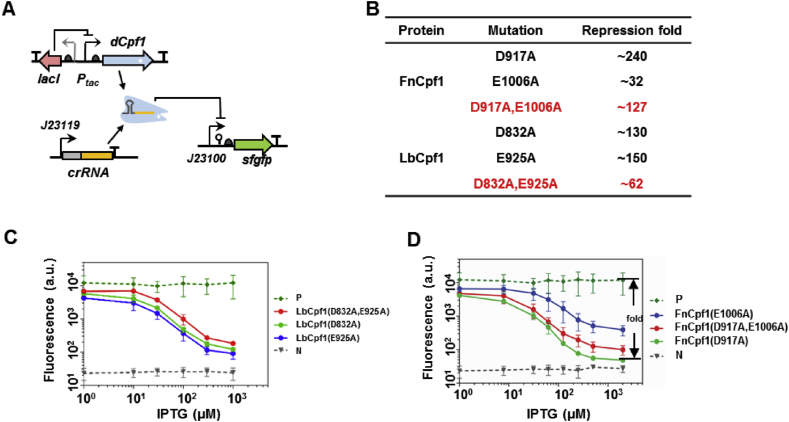
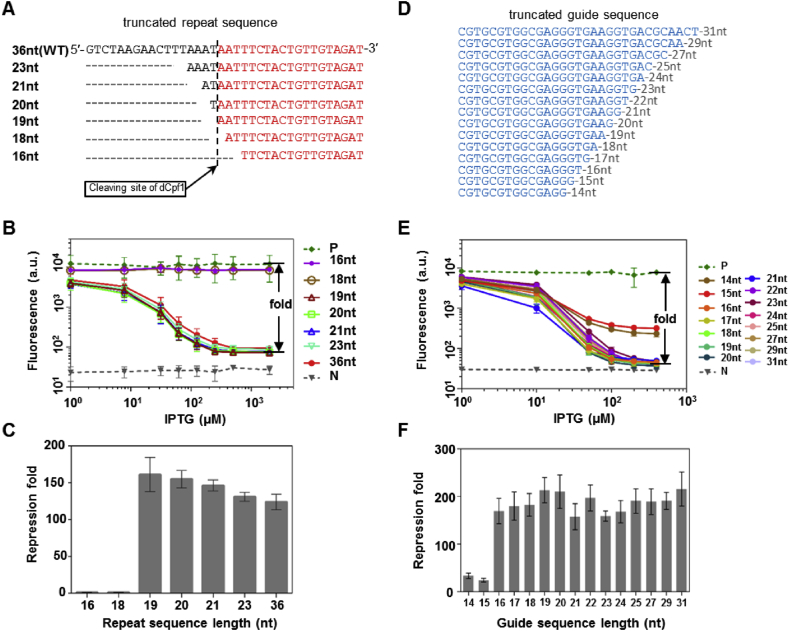
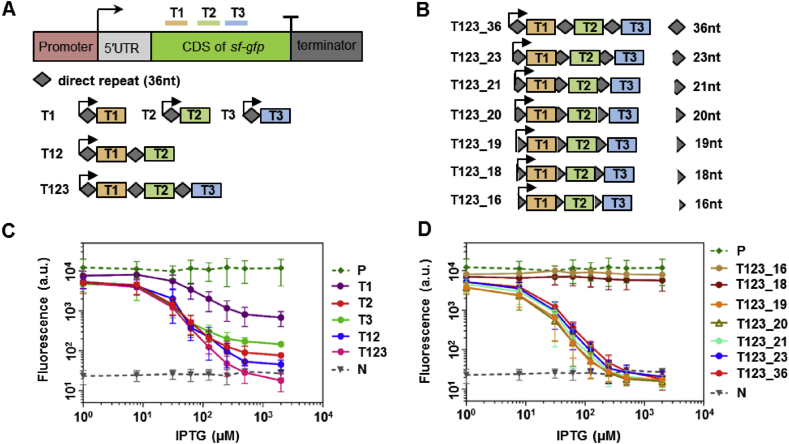
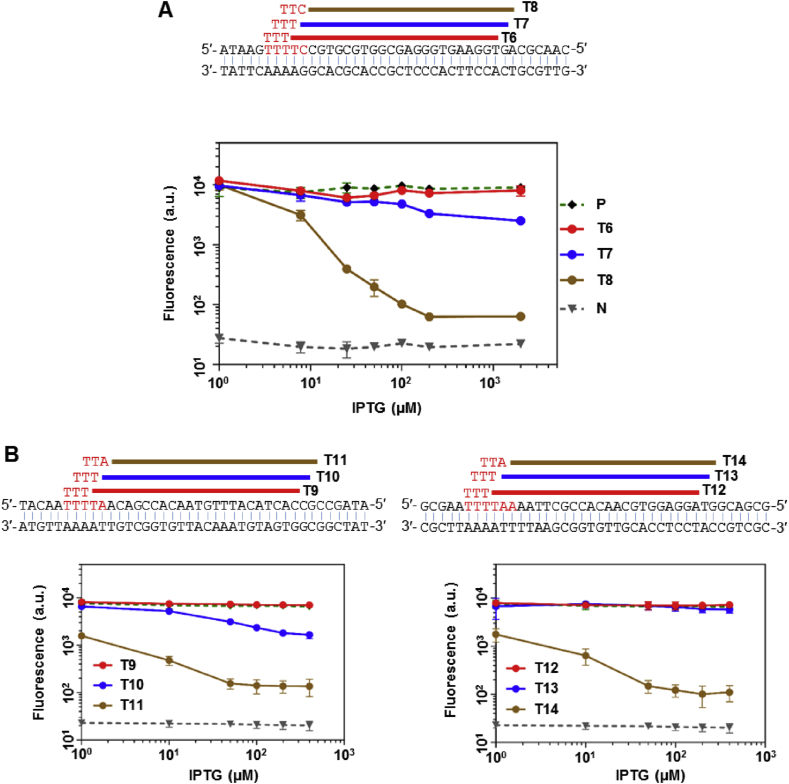
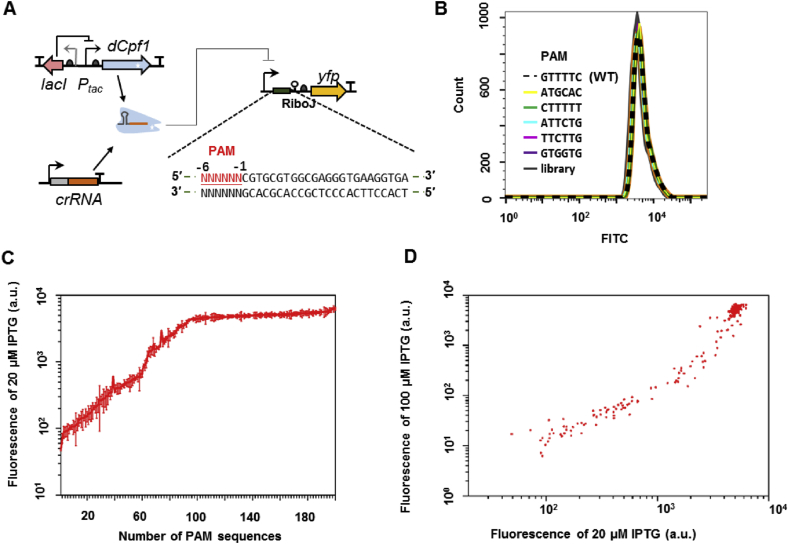
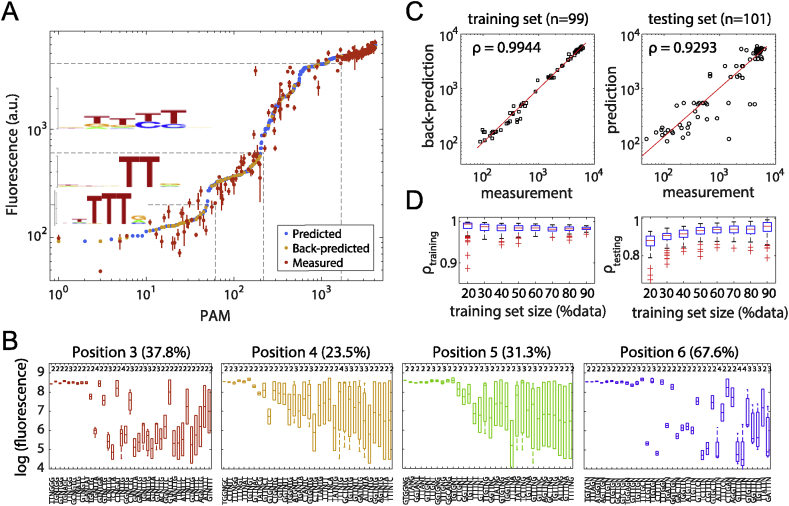
With a unique crRNA processing capability, the CRISPR associated Cpf1 protein holds great potential for multiplex gene regulation. Unlike the well-studied Cas9 protein, however, conversion of Cpf1 to a transcription regulator and its related properties have not been systematically explored yet. In this study, we investigated the mutation schemes and crRNA requirements for the DNase deactivated Cpf1 (dCpf1). By shortening the direct repeat sequence, we obtained genetically stable crRNA co-transcripts and improved gene repression with multiplex targeting. A screen of diversity-enriched PAM library was designed to investigate the PAM-dependency of gene regulation by dCpf1 from Francisella novicida and Lachnospiraceae bacterium. We found novel PAM patterns that elicited strong or medium gene repressions. Using a computational algorithm, we predicted regulatory outputs for all possible PAM sequences, which spanned a large dynamic range that could be leveraged for regulatory purposes. These newly identified features will facilitate the efficient design of CRISPR-dCpf1 based systems for tunable multiplex gene regulation.
Figures
References
-
- Cho S.W., Kim S., Kim J.M., Kim J.S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat Biotechnol. 2013;31:230–232. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
