

Peribiliary Glands Are Key in Regeneration of the Human Biliary Epithelium After Severe Bile Duct Injury
- PMID: 30506902
- PMCID: PMC6594148
- DOI: 10.1002/hep.30365
Peribiliary Glands Are Key in Regeneration of the Human Biliary Epithelium After Severe Bile Duct Injury
Abstract
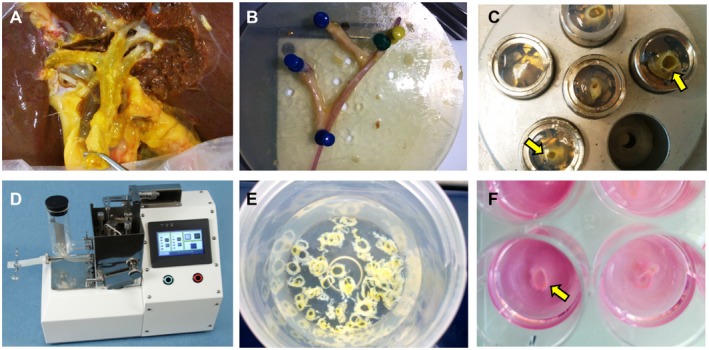
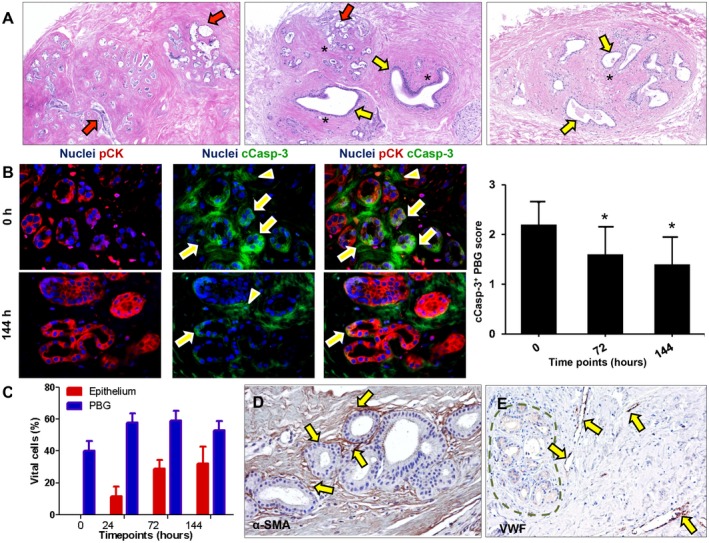
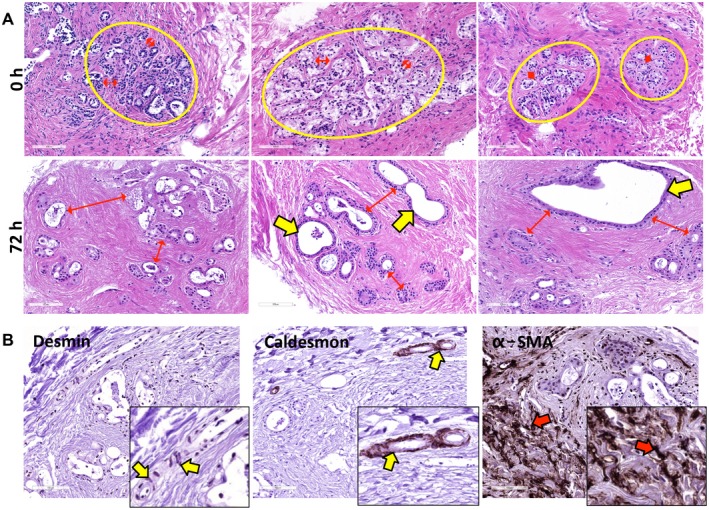
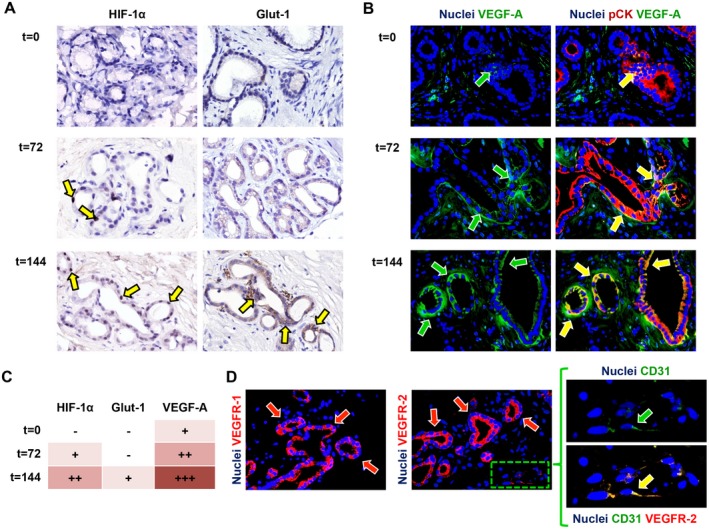
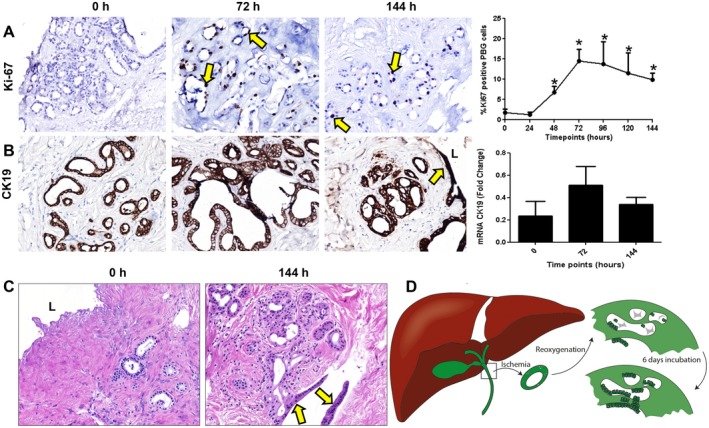
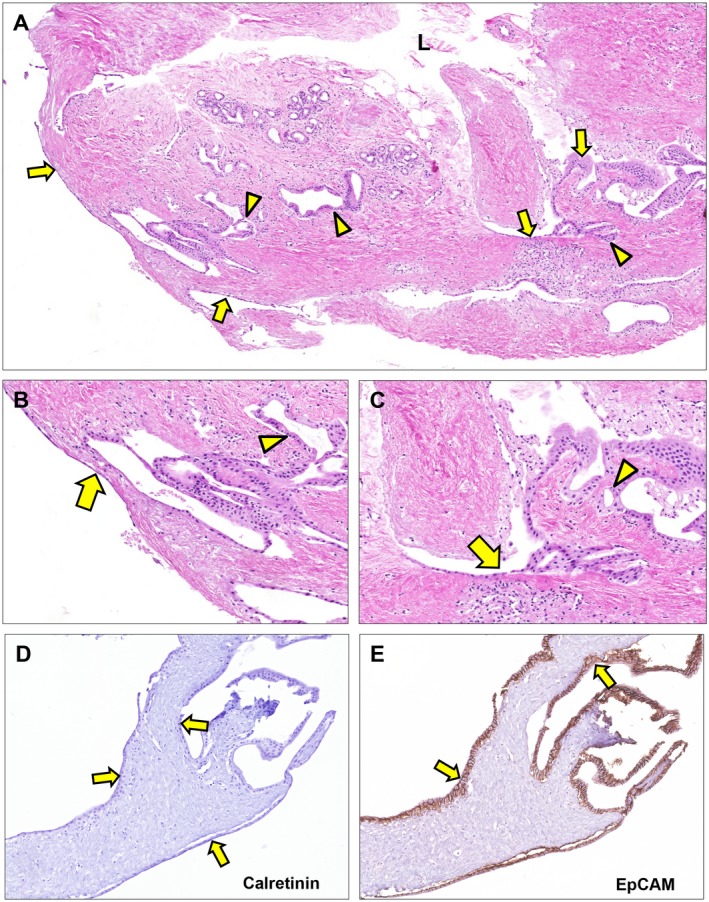
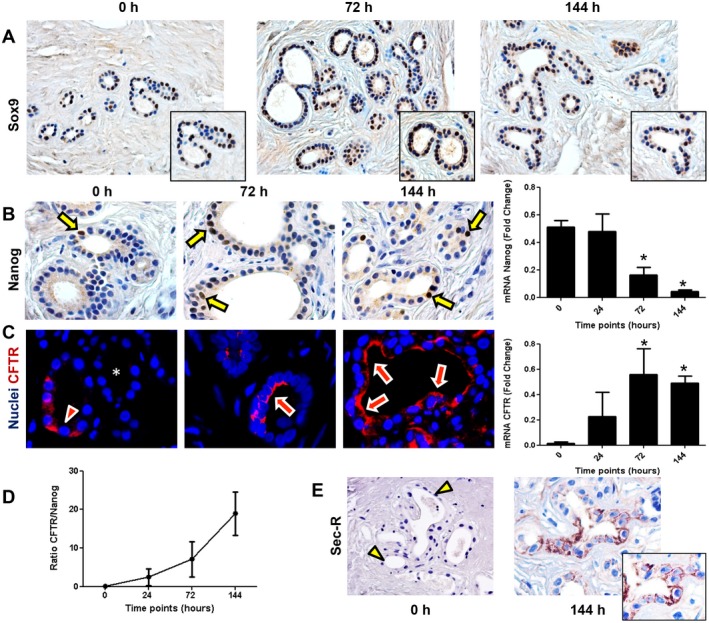
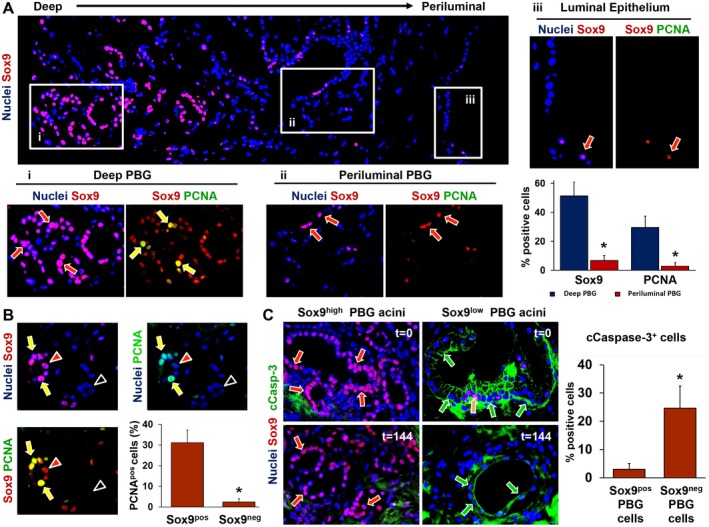
Peribiliary glands (PBG) are a source of stem/progenitor cells organized in a cellular network encircling large bile ducts. Severe cholangiopathy with loss of luminal biliary epithelium has been proposed to activate PBG, resulting in cell proliferation and differentiation to restore biliary epithelial integrity. However, formal evidence for this concept in human livers is lacking. We therefore developed an ex vivo model using precision-cut slices of extrahepatic human bile ducts obtained from discarded donor livers, providing an intact anatomical organization of cell structures, to study spatiotemporal differentiation and migration of PBG cells after severe biliary injury. Postischemic bile duct slices were incubated in oxygenated culture medium for up to a week. At baseline, severe tissue injury was evident with loss of luminal epithelial lining and mural stroma necrosis. In contrast, PBG remained relatively well preserved and different reactions of PBG were noted, including PBG dilatation, cell proliferation, and maturation. Proliferation of PBG cells increased after 24 hours of oxygenated incubation, reaching a peak after 72 hours. Proliferation of PBG cells was paralleled by a reduction in PBG apoptosis and differentiation from a primitive and pluripotent (homeobox protein Nanog+/ sex-determining region Y-box 9+) to a mature (cystic fibrosis transmembrane conductance regulator+/secretin receptor+) and activated phenotype (increased expression of hypoxia-inducible factor 1 alpha, glucose transporter 1, and vascular endothelial growth factor A). Migration of proliferating PBG cells in our ex vivo model was unorganized, but resulted in generation of epithelial monolayers at stromal surfaces. Conclusion: Human PBG contain biliary progenitor cells and are able to respond to bile duct epithelial loss with proliferation, differentiation, and maturation to restore epithelial integrity. The ex vivo spatiotemporal behavior of human PBG cells provides evidence for a pivotal role of PBG in biliary regeneration after severe injury.
© 2018 The Authors. Hepatology published by Wiley Periodicals, Inc., on behalf of American Association for the Study of Liver Diseases.
Figures
References
-
- Cardinale V, Wang Y, Carpino G, Cui CB, Gatto M, Rossi M, et al. Multipotent stem/progenitor cells in human biliary tree give rise to hepatocytes, cholangiocytes, and pancreatic islets. Hepatology 2011;54:2159‐2172. - PubMed
-
- Lanzoni G, Cardinale V, Carpino G. The hepatic, biliary, and pancreatic network of stem/progenitor cell niches in humans: A new reference frame for disease and regeneration. Hepatology 2016;64:277‐286. - PubMed
-
- de Jong IEM, van Leeuwen OB, Lisman T, Gouw ASH, Porte RJ. Repopulating the biliary tree from the peribiliary glands. Biochim Biophys Acta 2018;1864:1524‐1531. - PubMed
-
- Carpino G, Cardinale V, Renzi A, Hov JR, Berloco PB, Rossi M, et al. Activation of biliary tree stem cells within peribiliary glands in primary sclerosing cholangitis. J Hepatol 2015;63:1220‐1228. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
