

Bacterial sepsis triggers an antiviral response that causes translation shutdown
- PMID: 30507610
- PMCID: PMC6307966
- DOI: 10.1172/JCI123284
Bacterial sepsis triggers an antiviral response that causes translation shutdown
Abstract
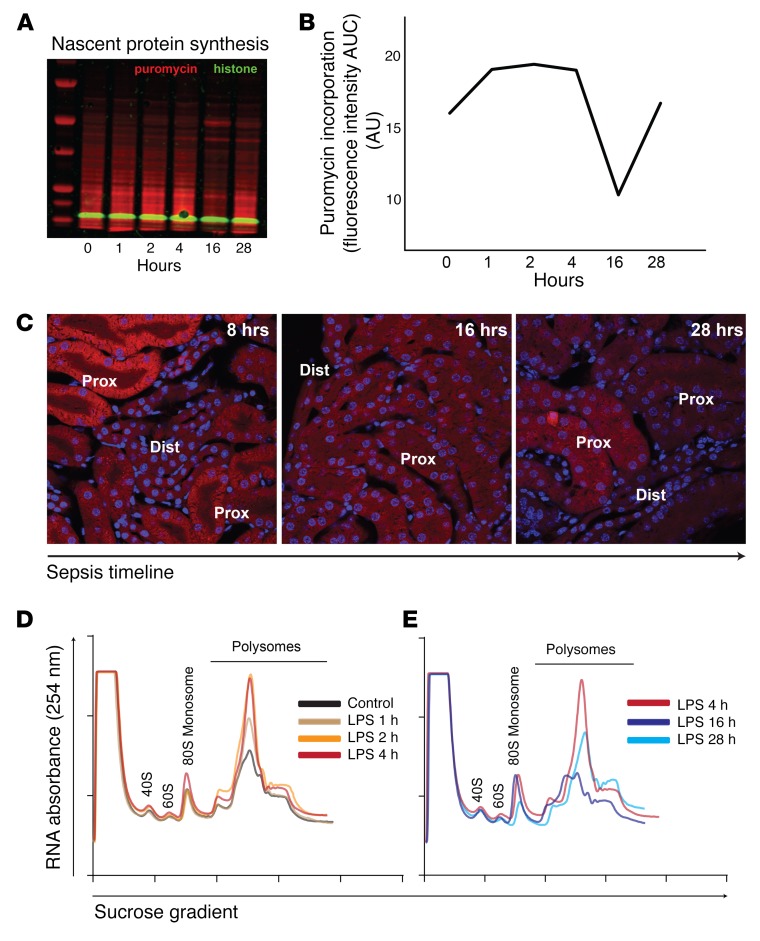
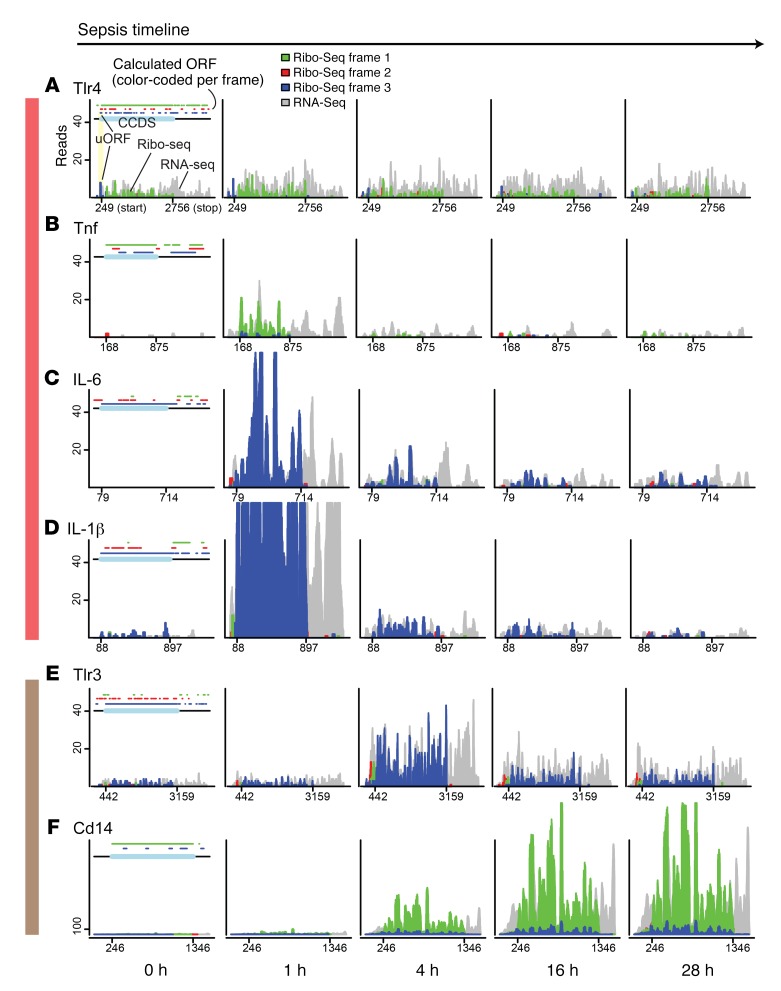
In response to viral pathogens, the host upregulates antiviral genes that suppress translation of viral mRNAs. However, induction of such antiviral responses may not be exclusive to viruses, as the pathways lie at the intersection of broad inflammatory networks that can also be induced by bacterial pathogens. Using a model of Gram-negative sepsis, we show that propagation of kidney damage initiated by a bacterial origin ultimately involves antiviral responses that result in host translation shutdown. We determined that activation of the eukaryotic translation initiation factor 2-α kinase 2/eukaryotic translation initiation factor 2α (Eif2ak2/Eif2α) axis is the key mediator of translation initiation block in late-phase sepsis. Reversal of this axis mitigated kidney injury. Furthermore, temporal profiling of the kidney translatome revealed that multiple genes involved in formation of the initiation complex were translationally altered during bacterial sepsis. Collectively, our findings imply that translation shutdown is indifferent to the specific initiating pathogen and is an important determinant of tissue injury in sepsis.
Keywords: Inflammation; Nephrology; Proteomics; Translation.
Conflict of interest statement
Figures







Comment in
-
Targeting protein translation to prevent septic kidney injury.J Clin Invest. 2019 Jan 2;129(1):60-62. doi: 10.1172/JCI125432. Epub 2018 Dec 3. J Clin Invest. 2019. PMID: 30507607 Free PMC article.
-
Sepsis halts renal protein translation.Nat Rev Nephrol. 2019 Mar;15(3):124. doi: 10.1038/s41581-018-0104-5. Nat Rev Nephrol. 2019. PMID: 30563968 No abstract available.
References
-
- Angus DC, van der Poll T. Severe sepsis and septic shock. N Engl J Med. 2013;369(21):2063. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
