

Tissue tropisms opt for transmissible reassortants during avian and swine influenza A virus co-infection in swine
- PMID: 30507946
- PMCID: PMC6292640
- DOI: 10.1371/journal.ppat.1007417
Tissue tropisms opt for transmissible reassortants during avian and swine influenza A virus co-infection in swine
Abstract
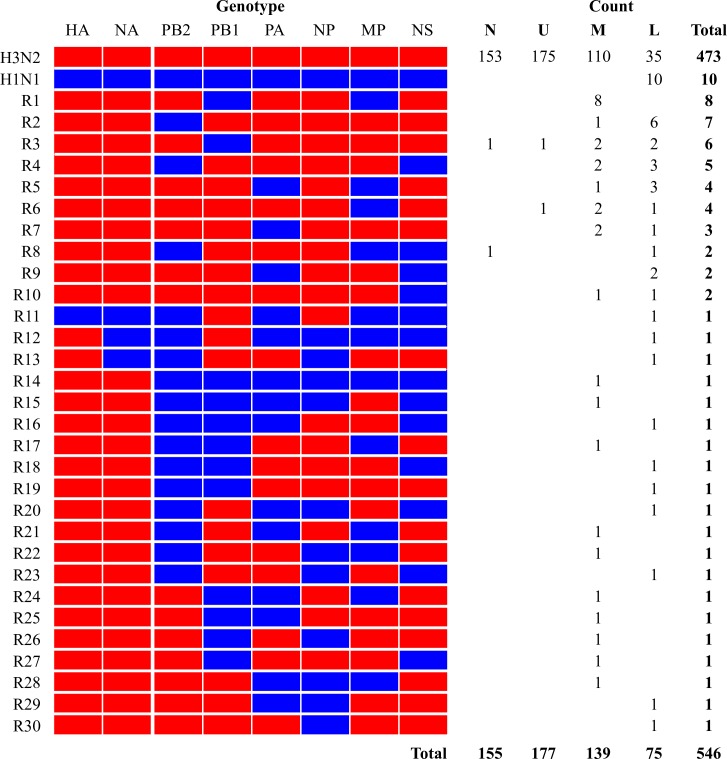
Genetic reassortment between influenza A viruses (IAVs) facilitate emergence of pandemic strains, and swine are proposed as a "mixing vessel" for generating reassortants of avian and mammalian IAVs that could be of risk to mammals, including humans. However, how a transmissible reassortant emerges in swine are not well understood. Genomic analyses of 571 isolates recovered from nasal wash samples and respiratory tract tissues of a group of co-housed pigs (influenza-seronegative, avian H1N1 IAV-infected, and swine H3N2 IAV-infected pigs) identified 30 distinct genotypes of reassortants. Viruses recovered from lower respiratory tract tissues had the largest genomic diversity, and those recovered from turbinates and nasal wash fluids had the least. Reassortants from lower respiratory tracts had the largest variations in growth kinetics in respiratory tract epithelial cells, and the cold temperature in swine nasal cells seemed to select the type of reassortant viruses shed by the pigs. One reassortant in nasal wash samples was consistently identified in upper, middle, and lower respiratory tract tissues, and it was confirmed to be transmitted efficiently between pigs. Study findings suggest that, during mixed infections of avian and swine IAVs, genetic reassortments are likely to occur in the lower respiratory track, and tissue tropism is an important factor selecting for a transmissible reassortant.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Tong S, Li Y, Rivailler P, Conrardy C, Castillo DA, Chen LM, et al. A distinct lineage of influenza A virus from bats. Proc Natl Acad Sci U S A. 2012;109(11):4269–74. 10.1073/pnas.1116200109 ; PubMed Central PMCID: PMCPMC3306675. - DOI - PMC - PubMed
-
- Tong S, Zhu X, Li Y, Shi M, Zhang J, Bourgeois M, et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013;9(10):e1003657 10.1371/journal.ppat.1003657 ; PubMed Central PMCID: PMCPMC3794996. - DOI - PMC - PubMed
-
- Olsen B, Munster VJ, Wallensten A, Waldenstrom J, Osterhaus AD, Fouchier RA. Global patterns of influenza a virus in wild birds. Science. 2006;312(5772):384–8. 10.1126/science.1122438 . - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
