

Impaired male fertility and abnormal epididymal epithelium differentiation in mice lacking CRISP1 and CRISP4
- PMID: 30510210
- PMCID: PMC6277452
- DOI: 10.1038/s41598-018-35719-3
Impaired male fertility and abnormal epididymal epithelium differentiation in mice lacking CRISP1 and CRISP4
Abstract
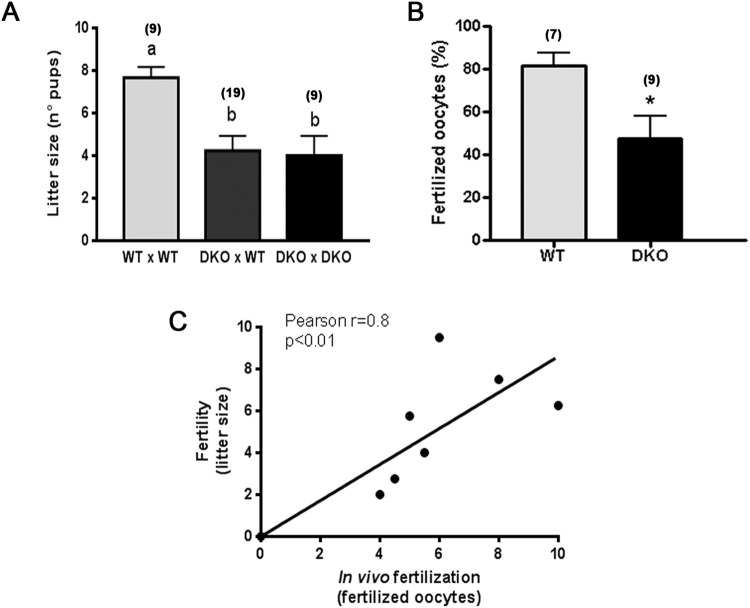
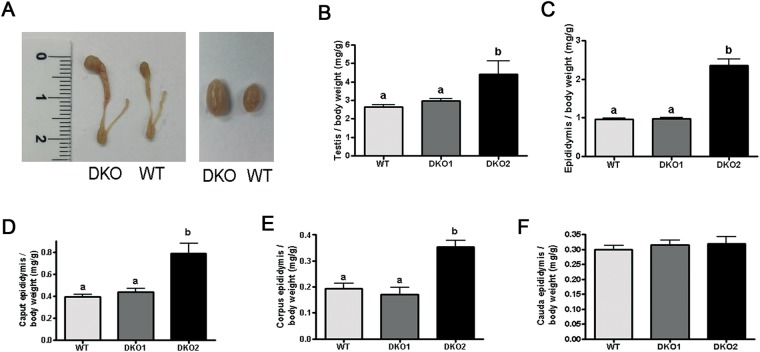
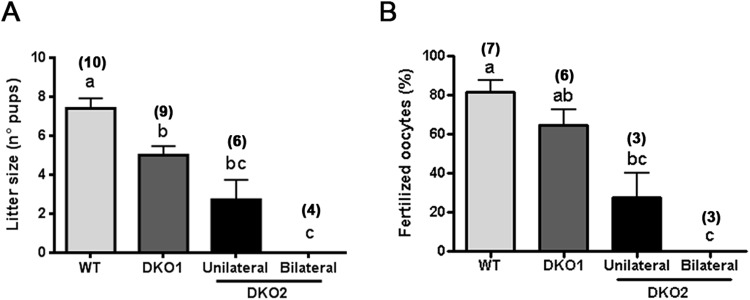
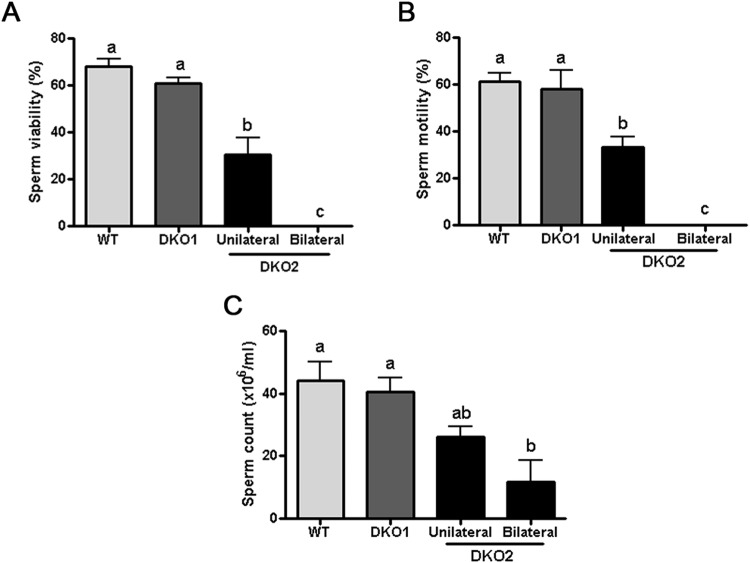
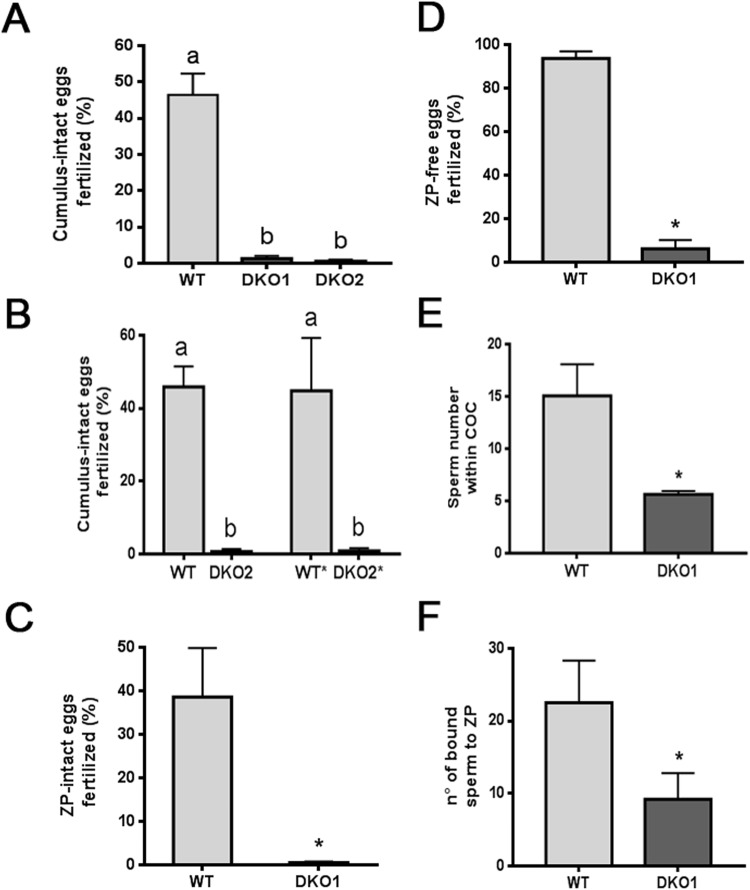
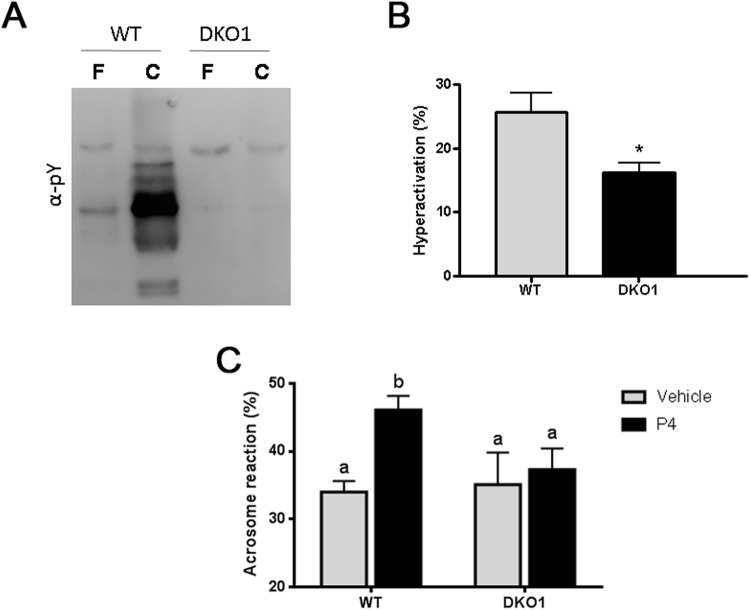
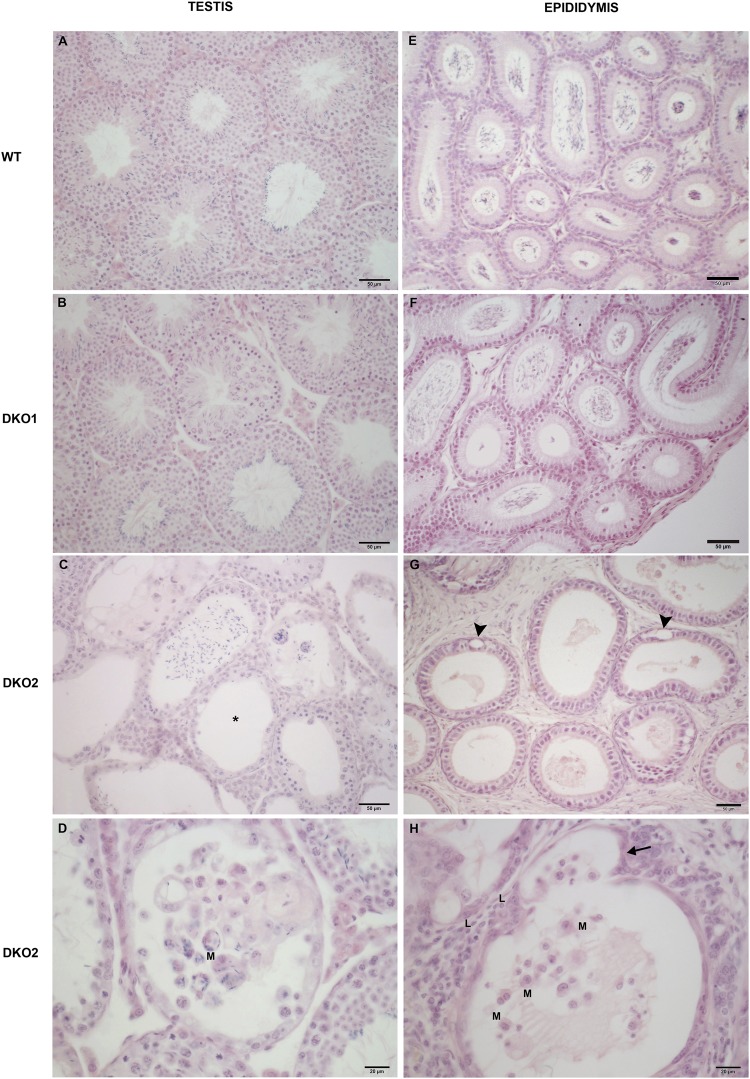
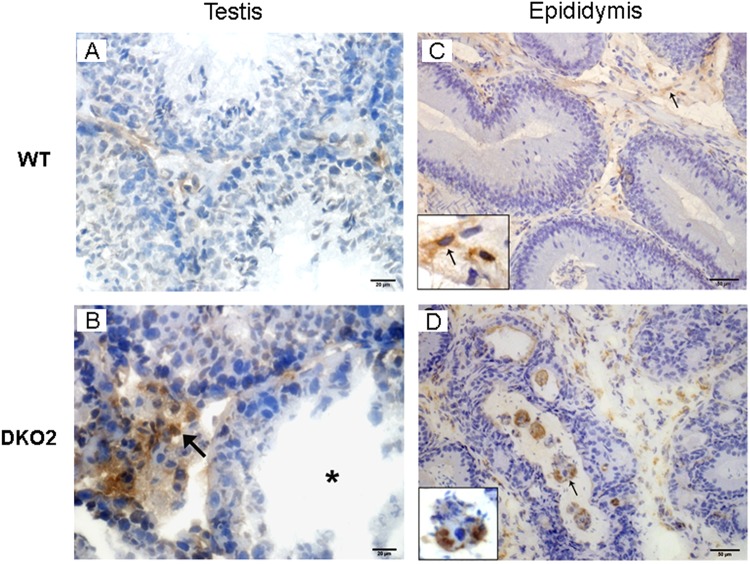
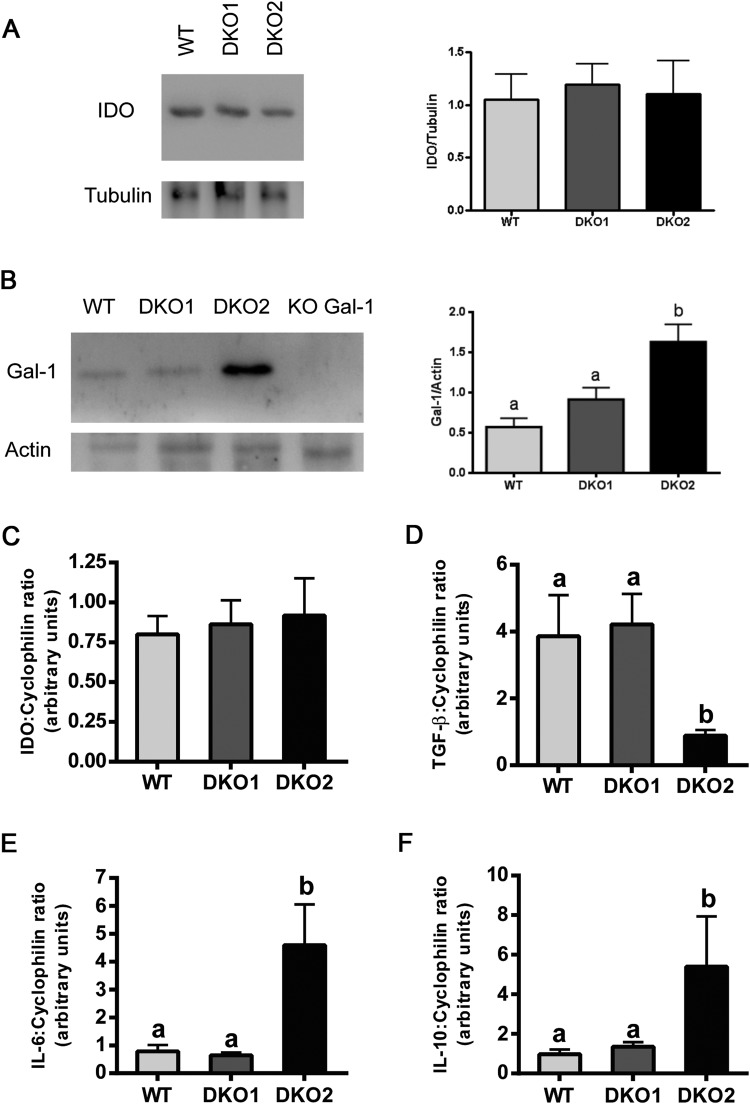
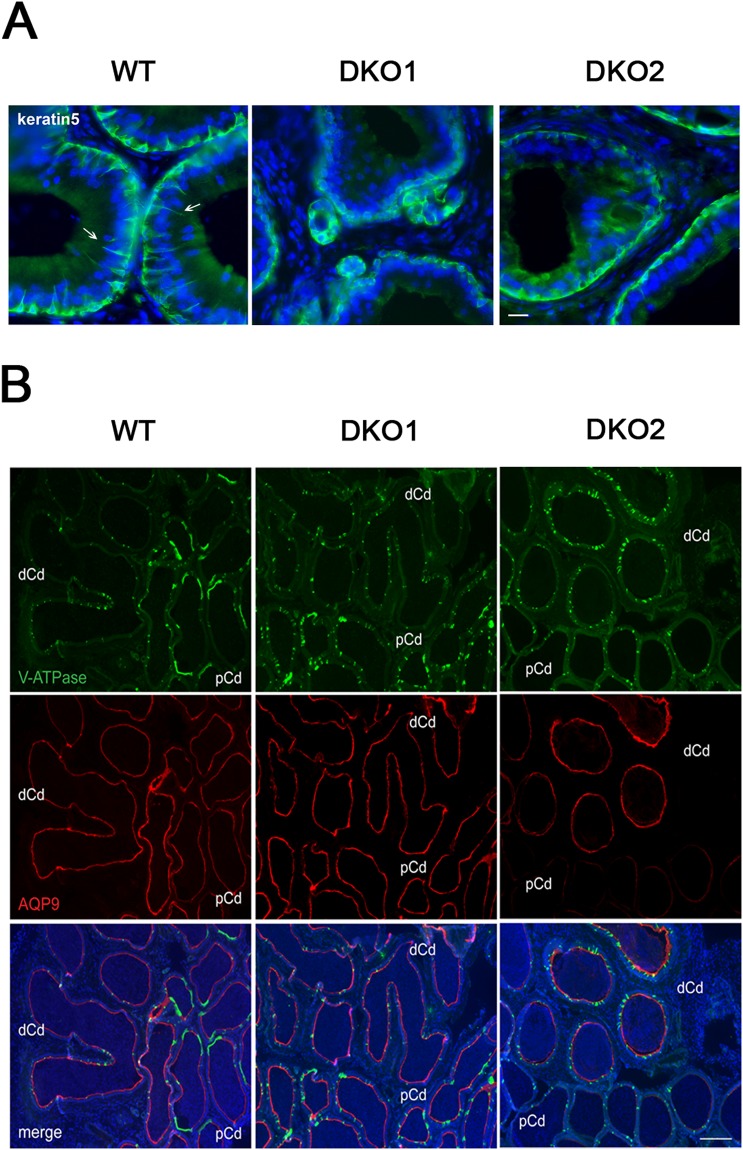
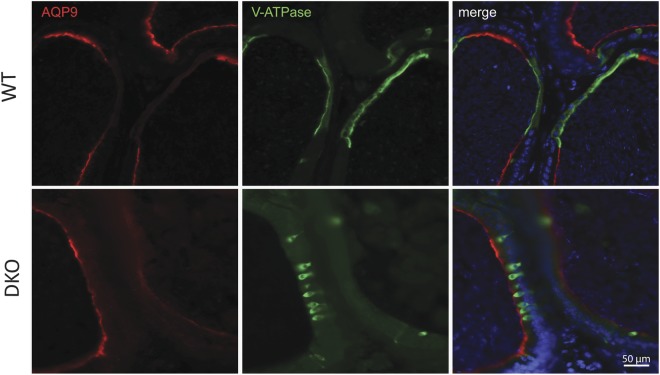
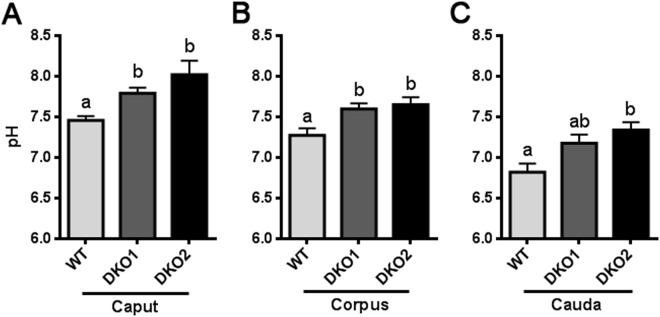
Epididymal Cysteine Rich Secretory Proteins 1 and 4 (CRISP1 and CRISP4) associate with sperm during maturation and play different roles in fertilization. However, males lacking each of these molecules individually are fertile, suggesting compensatory mechanisms between these homologous proteins. Based on this, in the present work, we generated double CRISP1/CRISP4 knockout (DKO) mice and examined their reproductive phenotype. Our data showed that the simultaneous lack of the two epididymal proteins results in clear fertility defects. Interestingly, whereas most of the animals exhibited specific sperm fertilizing ability defects supportive of the role of CRISP proteins in fertilization, one third of the males showed an unexpected epididymo-orchitis phenotype with altered levels of inflammatory molecules and non-viable sperm in the epididymis. Further analysis showed that DKO mice exhibited an immature epididymal epithelium and abnormal luminal pH, supporting these defects as likely responsible for the different phenotypes observed. These observations reveal that CRISP proteins are relevant for epididymal epithelium differentiation and male fertility, contributing to a better understanding of the fine-tuning mechanisms underlying sperm maturation and immunotolerance in the epididymis with clear implications for human epididymal physiology and pathology.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Yanagimachi, R. In The Physiologyof Reproduction (eds Knobil, E. & Neill, J. D.) 189–317 (1994).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
