

Amyloid-like Behavior of Site-Specifically Citrullinated Myelin Oligodendrocyte Protein (MOG) Peptide Fragments inside EBV-Infected B-Cells Influences Their Cytotoxicity and Autoimmunogenicity
- PMID: 30513201
- PMCID: PMC6374747
- DOI: 10.1021/acs.biochem.8b00852
Amyloid-like Behavior of Site-Specifically Citrullinated Myelin Oligodendrocyte Protein (MOG) Peptide Fragments inside EBV-Infected B-Cells Influences Their Cytotoxicity and Autoimmunogenicity
Abstract
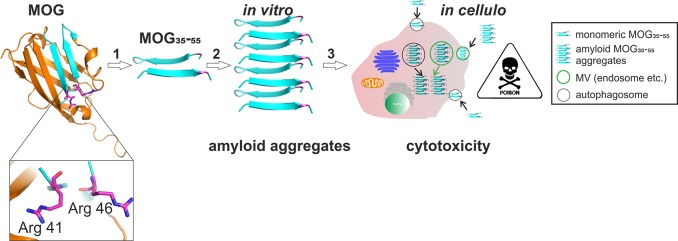
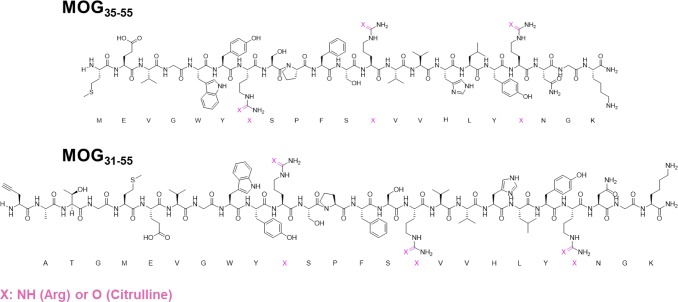
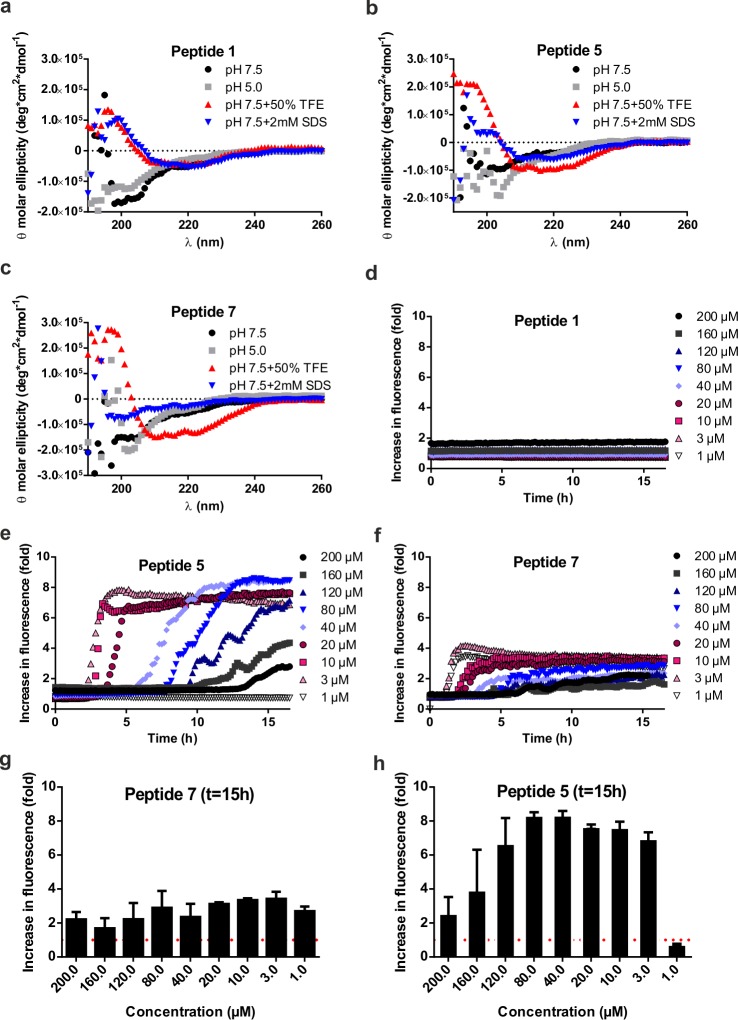
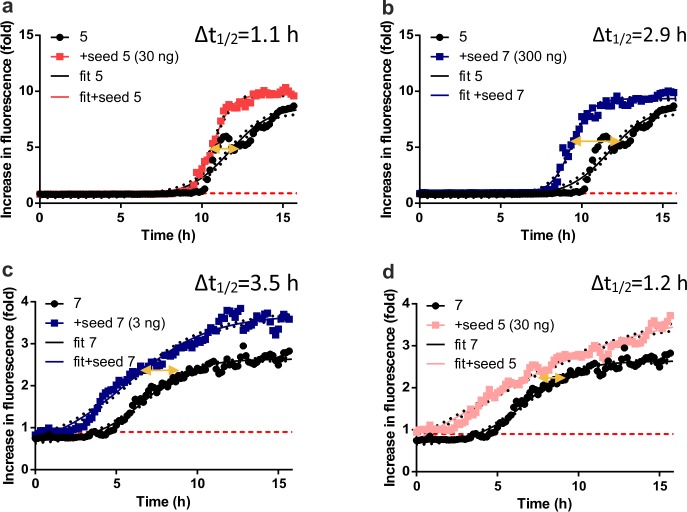
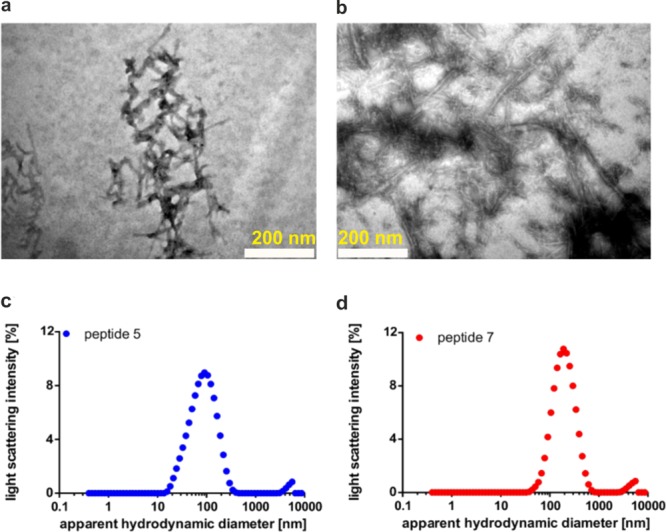
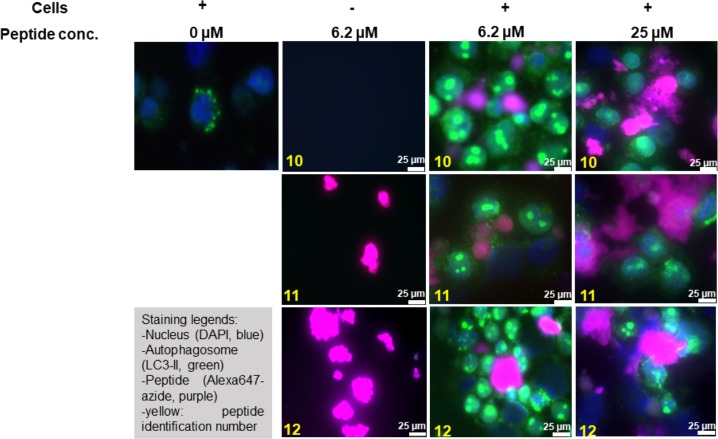
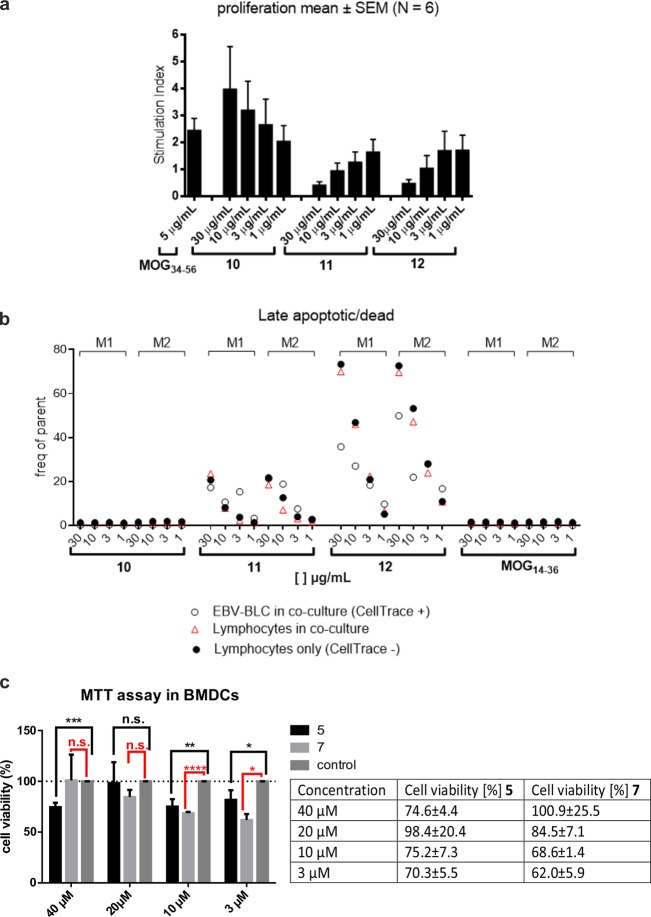
Multiple sclerosis (MS) is an autoimmune disorder manifested via chronic inflammation, demyelination, and neurodegeneration inside the central nervous system. The progressive phase of MS is characterized by neurodegeneration, but unlike classical neurodegenerative diseases, amyloid-like aggregation of self-proteins has not been documented. There is evidence that citrullination protects an immunodominant peptide of human myelin oligodendrocyte glycoprotein (MOG34-56) against destructive processing in Epstein-Barr virus-infected B-lymphocytes (EBV-BLCs) in marmosets and causes exacerbation of ongoing MS-like encephalopathies in mice. Here we collected evidence that citrullination of MOG can also lead to amyloid-like behavior shifting the disease pathogenesis toward neurodegeneration. We observed that an immunodominant MOG peptide, MOG35-55, displays amyloid-like behavior upon site-specific citrullination at positions 41, 46, and/or 52. These amyloid aggregates are shown to be toxic to the EBV-BLCs and to dendritic cells at concentrations favored for antigen presentation, suggesting a role of amyloid-like aggregation in the pathogenesis of progressive MS.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Lublin F. D.; Reingold S. C.; Cohen J. A.; Cutter G. R.; Sørensen P. S.; Thompson A. J.; Wolinsky J. S.; Balcer L. J.; Banwell B.; Barkhof F.; Bebo B.; Calabresi P. A.; Clanet M.; Comi G.; Fox R. J.; Freedman M. S.; Goodman A. D.; Inglese M.; Kappos L.; Kieseier B. C.; Lincoln J. A.; Lubetzki C.; Miller A. E.; Montalban X.; O’Connor P. W.; Petkau J.; Pozzilli C.; Rudick R. A.; Sormani M. P.; Stüve O.; Waubant E.; Polman C. H. (2014) Defining the clinical course of multiple sclerosis. Neurology 83 (3), 278–286. 10.1212/WNL.0000000000000560. - DOI - PMC - PubMed
-
- Sumaya C. V.; Myers L.; Ellison G. W. (1976) Epstein-Barr virus antibodies in multiple sclerosis. Trans. Am. Neurol. Assoc. 101, 300. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
