

Downregulation of RdDM during strawberry fruit ripening
- PMID: 30514401
- PMCID: PMC6280534
- DOI: 10.1186/s13059-018-1587-x
Downregulation of RdDM during strawberry fruit ripening
Abstract
Background: Recently, DNA methylation was proposed to regulate fleshy fruit ripening. Fleshy fruits can be distinguished by their ripening process as climacteric fruits, such as tomatoes, or non-climacteric fruits, such as strawberries. Tomatoes undergo a global decrease in DNA methylation during ripening, due to increased expression of a DNA demethylase gene. The dynamics and biological relevance of DNA methylation during the ripening of non-climacteric fruits are unknown.
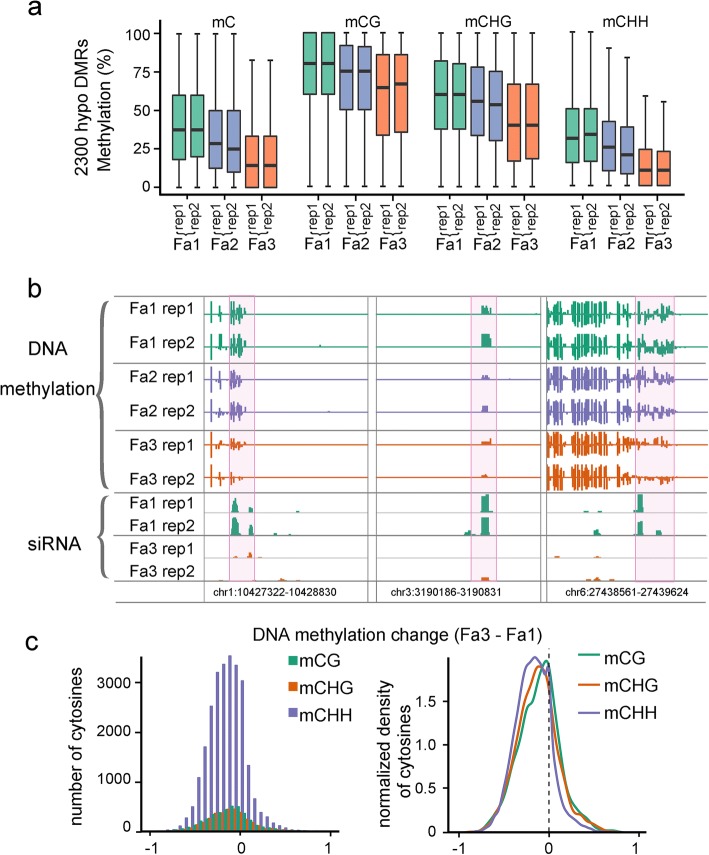
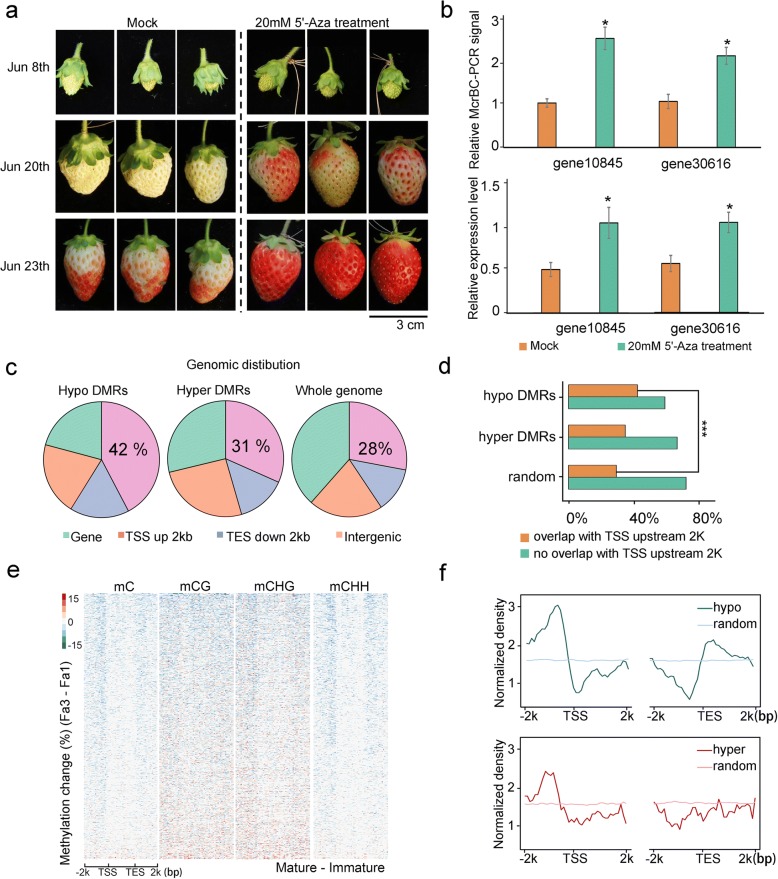
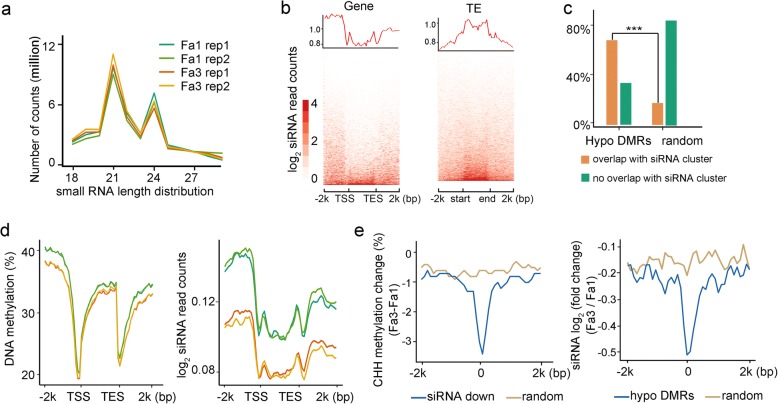
Results: Here, we generate single-base resolution maps of the DNA methylome in immature and ripe strawberry. We observe an overall loss of DNA methylation during strawberry fruit ripening. Thus, ripening-induced DNA hypomethylation occurs not only in climacteric fruit, but also in non-climacteric fruit. Application of a DNA methylation inhibitor causes an early ripening phenotype, suggesting that DNA hypomethylation is important for strawberry fruit ripening. The mechanisms underlying DNA hypomethylation during the ripening of tomato and strawberry are distinct. Unlike in tomatoes, DNA demethylase genes are not upregulated during the ripening of strawberries. Instead, genes involved in RNA-directed DNA methylation are downregulated during strawberry ripening. Further, ripening-induced DNA hypomethylation is associated with decreased siRNA levels, consistent with reduced RdDM activity. Therefore, we propose that a downregulation of RdDM contributes to DNA hypomethylation during strawberry ripening.
Conclusions: Our findings provide new insight into the DNA methylation dynamics during the ripening of non-climacteric fruit and suggest a novel function of RdDM in regulating an important process in plant development.
Keywords: DNA methylation; Fruit ripening; RdDM; Strawberry; siRNA.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures



References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
