

TUBA1A mutations identified in lissencephaly patients dominantly disrupt neuronal migration and impair dynein activity
- PMID: 30517687
- PMCID: PMC6452179
- DOI: 10.1093/hmg/ddy416
TUBA1A mutations identified in lissencephaly patients dominantly disrupt neuronal migration and impair dynein activity
Abstract
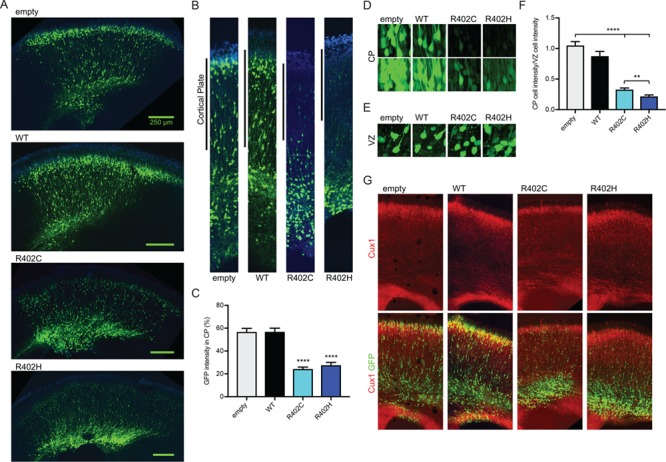
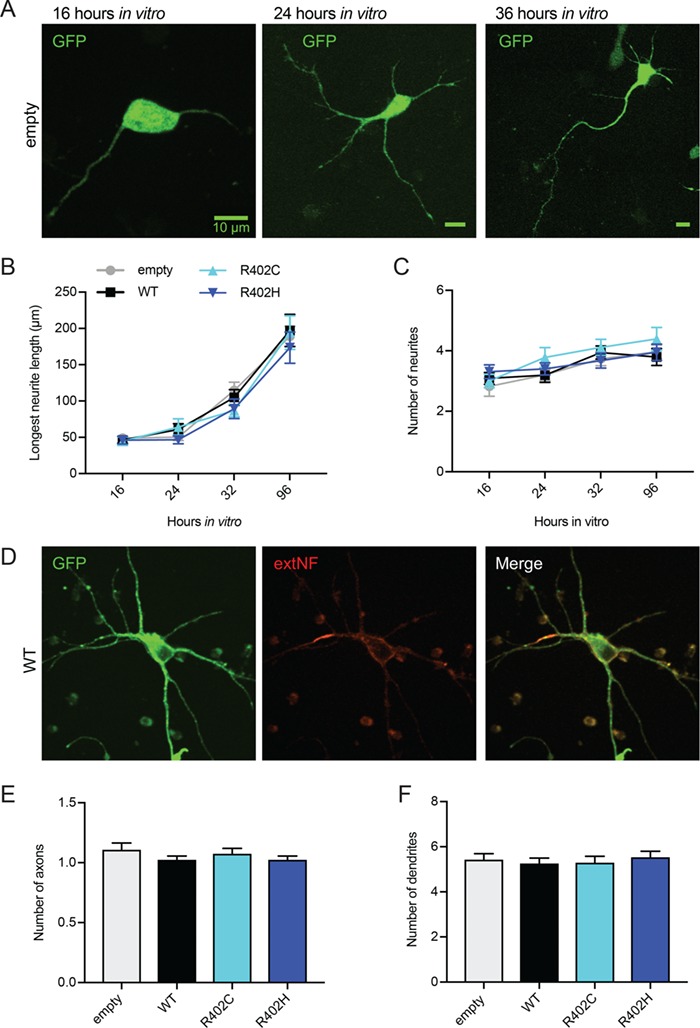
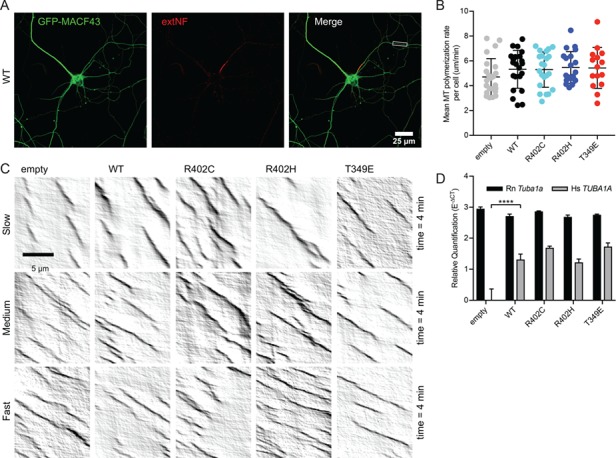
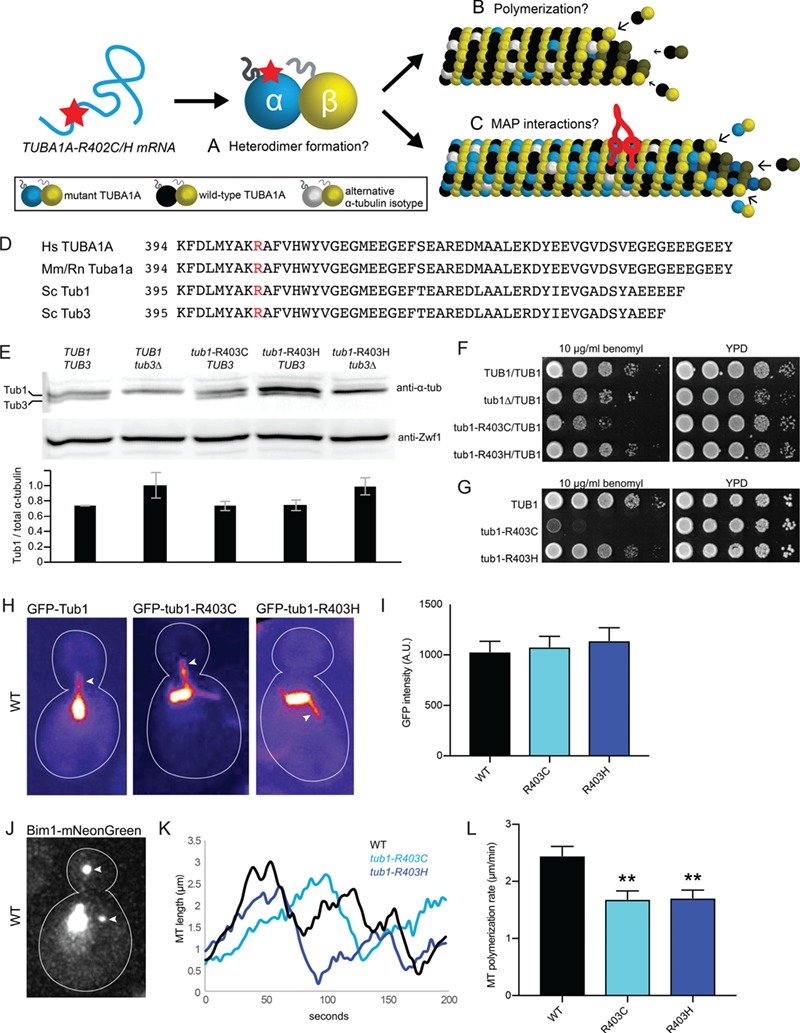
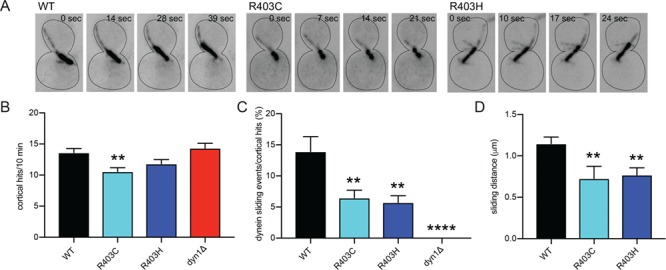
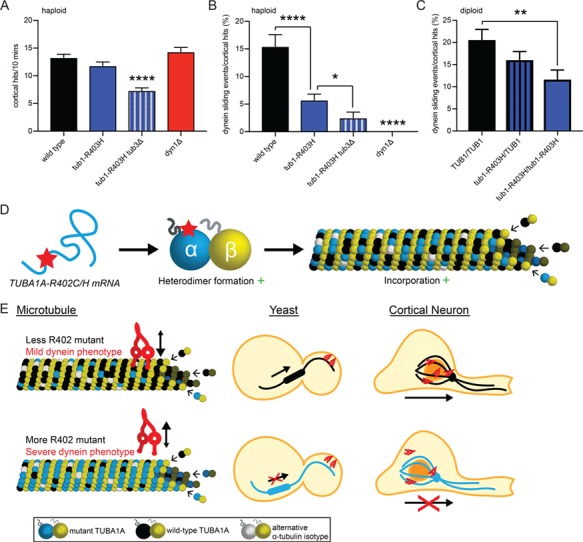
The microtubule cytoskeleton supports diverse cellular morphogenesis and migration processes during brain development. Mutations in tubulin genes are associated with severe human brain malformations known as 'tubulinopathies'; however, it is not understood how molecular-level changes in microtubule subunits lead to brain malformations. In this study, we demonstrate that missense mutations affecting arginine at position 402 (R402) of TUBA1A α-tubulin selectively impair dynein motor activity and severely and dominantly disrupt cortical neuronal migration. TUBA1A is the most commonly affected tubulin gene in tubulinopathy patients, and mutations altering R402 account for 30% of all reported TUBA1A mutations. We show for the first time that ectopic expression of TUBA1A-R402C and TUBA1A-R402H patient alleles is sufficient to dominantly disrupt cortical neuronal migration in the developing mouse brain, strongly supporting a causal role in the pathology of brain malformation. To isolate the precise molecular impact of R402 mutations, we generated analogous R402C and R402H mutations in budding yeast α-tubulin, which exhibit a simplified microtubule cytoskeleton. We find that R402 mutant tubulins assemble into microtubules that support normal kinesin motor activity but fail to support the activity of dynein motors. Importantly, the level of dynein impairment scales with the expression level of the mutant in the cell, suggesting a 'poisoning' mechanism in which R402 mutant α-tubulin acts dominantly by populating microtubules with defective binding sites for dynein. Based on our results, we propose a new model for the molecular pathology of tubulinopathies that may also extend to other tubulin-related neuropathies.
© The Author(s) 2018. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Bahi-Buisson N., Poirier K., Fourniol F., Saillour Y., Valence S., Lebrun N., Hully M., Bianco C.F., Boddaert N., Elie C. et al. (2014) The wide spectrum of tubulinopathies: what are the key features for the diagnosis. Brain, 137, 1676–1700. - PubMed
-
- Fallet-Bianco C., Laquerrière A., Poirier K., Razavi F., Guimiot F., Dias P., Loeuillet L., Lascelles K., Beldjord C., Carion N. et al. (2014) Mutations in tubulin genes are frequent causes of various foetal malformations of cortical development including microlissencephaly. Acta Neuropathol. Commun., 2, 69. - PMC - PubMed
-
- Coksaygan T., Magnus T., Cai J., Mughal M., Lepore A., Xue H., Fischer I. and Rao M.S. (2006) Neurogenesis in Talpha-1 tubulin transgenic mice during development and after injury. Exp. Neurol., 197, 475–485. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
