

NmeCas9 is an intrinsically high-fidelity genome-editing platform
- PMID: 30518407
- PMCID: PMC6282386
- DOI: 10.1186/s13059-018-1591-1
NmeCas9 is an intrinsically high-fidelity genome-editing platform
Abstract
Background: The development of CRISPR genome editing has transformed biomedical research. Most applications reported thus far rely upon the Cas9 protein from Streptococcus pyogenes SF370 (SpyCas9). With many RNA guides, wildtype SpyCas9 can induce significant levels of unintended mutations at near-cognate sites, necessitating substantial efforts toward the development of strategies to minimize off-target activity. Although the genome-editing potential of thousands of other Cas9 orthologs remains largely untapped, it is not known how many will require similarly extensive engineering to achieve single-site accuracy within large genomes. In addition to its off-targeting propensity, SpyCas9 is encoded by a relatively large open reading frame, limiting its utility in applications that require size-restricted delivery strategies such as adeno-associated virus vectors. In contrast, some genome-editing-validated Cas9 orthologs are considerably smaller and therefore better suited for viral delivery.
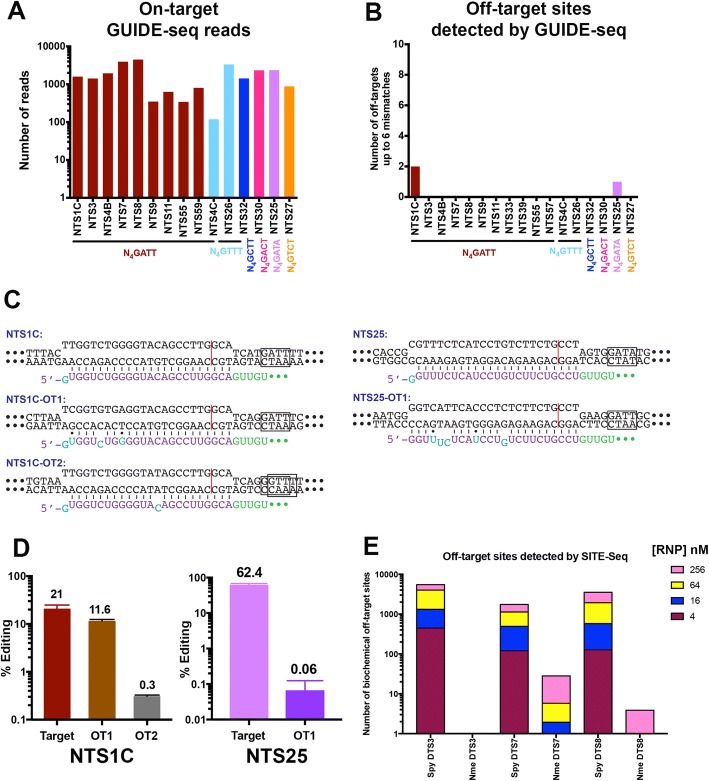
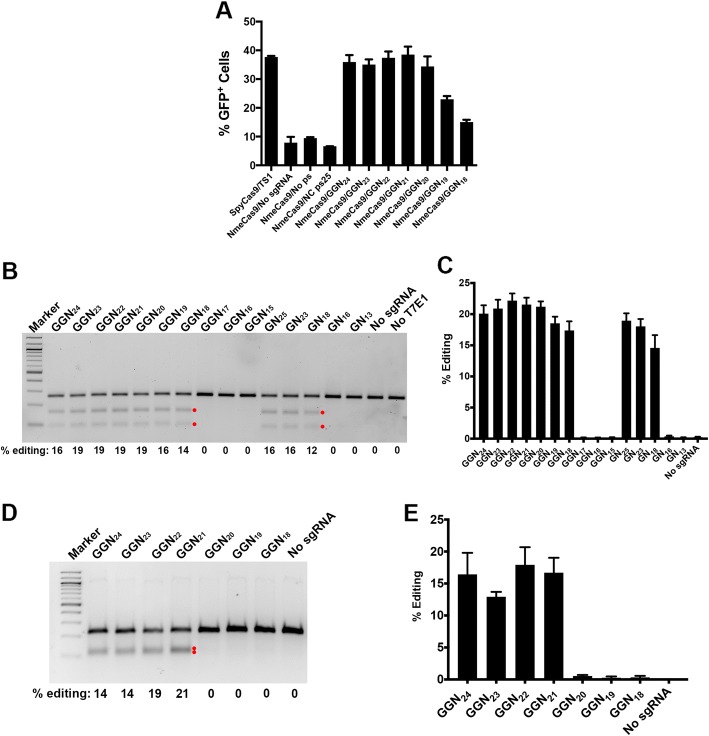
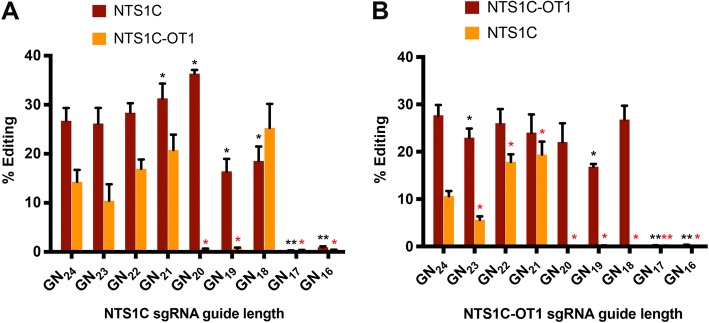
Results: Here we show that wildtype NmeCas9, when programmed with guide sequences of the natural length of 24 nucleotides, exhibits a nearly complete absence of unintended editing in human cells, even when targeting sites that are prone to off-target activity with wildtype SpyCas9. We also validate at least six variant protospacer adjacent motifs (PAMs), in addition to the preferred consensus PAM (5'-N4GATT-3'), for NmeCas9 genome editing in human cells.
Conclusions: Our results show that NmeCas9 is a naturally high-fidelity genome-editing enzyme and suggest that additional Cas9 orthologs may prove to exhibit similarly high accuracy, even without extensive engineering.
Keywords: CRISPR; Cas9; Neisseria meningitidis; Off-target; Protospacer adjacent motif; sgRNA.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
E.J.S. is a co-founder and scientific advisor of Intellia Therapeutics. P.D.D, A.H.S, A.M.L, K.M, C.K.F, and P.C are current or former employees of Caribou Biosciences, Inc., a company that develops and commercializes genome engineering technologies, and such individuals may own shares or stock options in Caribou Biosciences. SITE-Seq is a trademark of Caribou Biosciences, Inc. The other authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
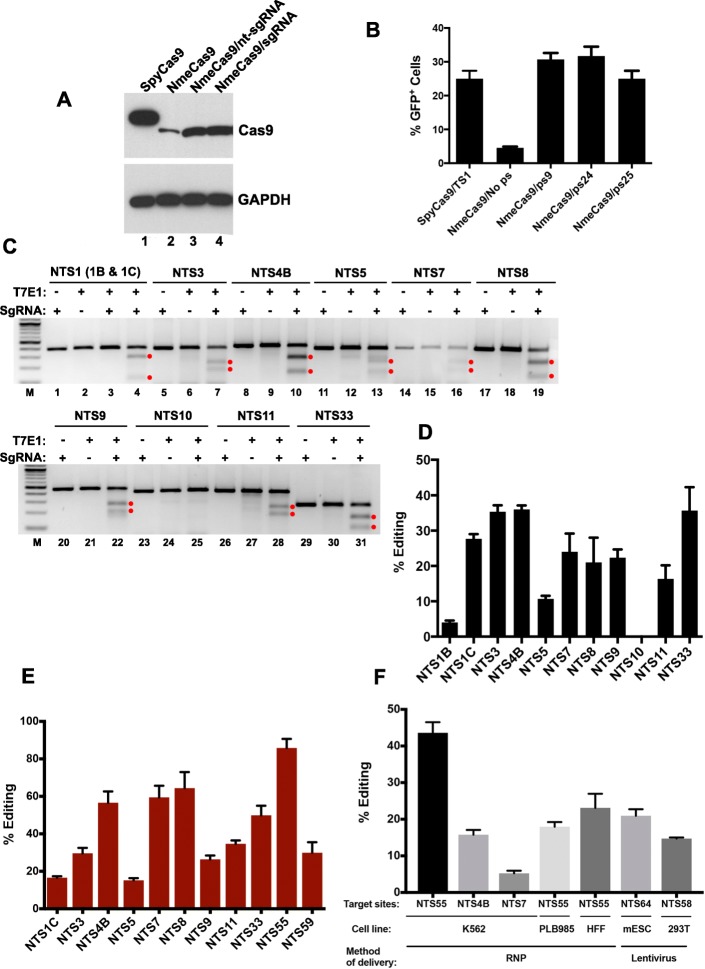
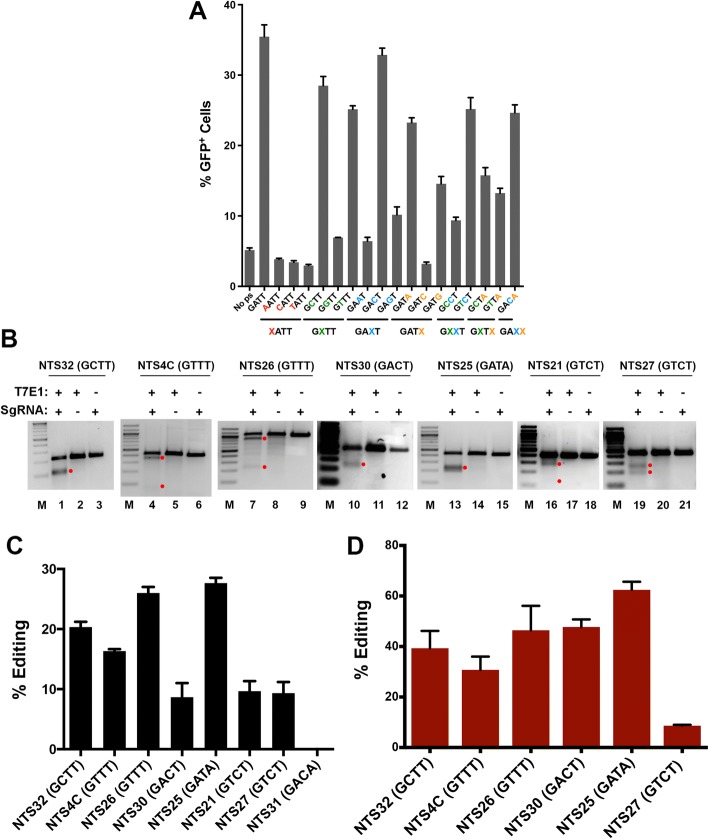
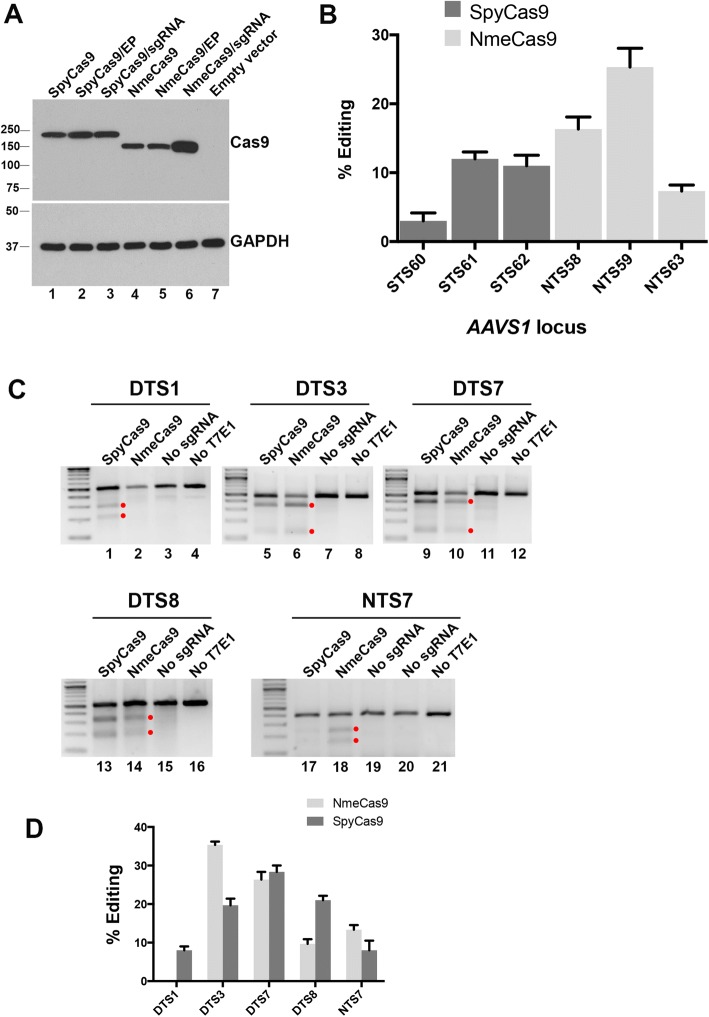
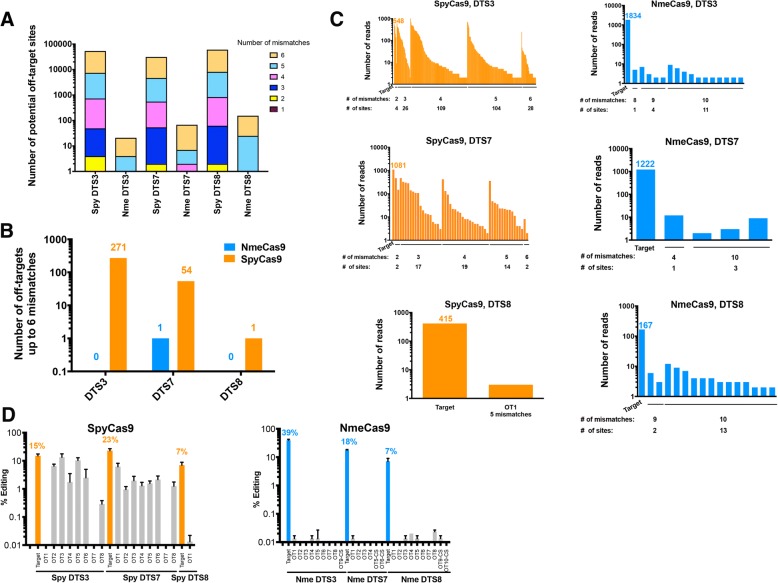
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
