

A common transcriptomic program acquired in the thymus defines tissue residency of MAIT and NKT subsets
- PMID: 30518599
- PMCID: PMC6314520
- DOI: 10.1084/jem.20181483
A common transcriptomic program acquired in the thymus defines tissue residency of MAIT and NKT subsets
Abstract
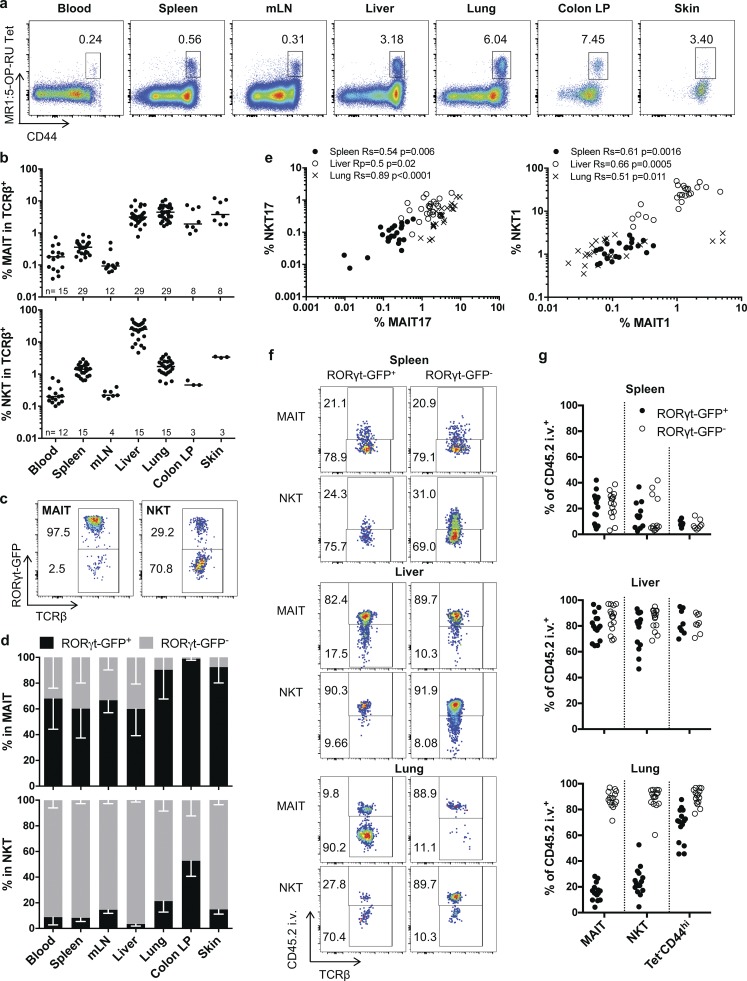
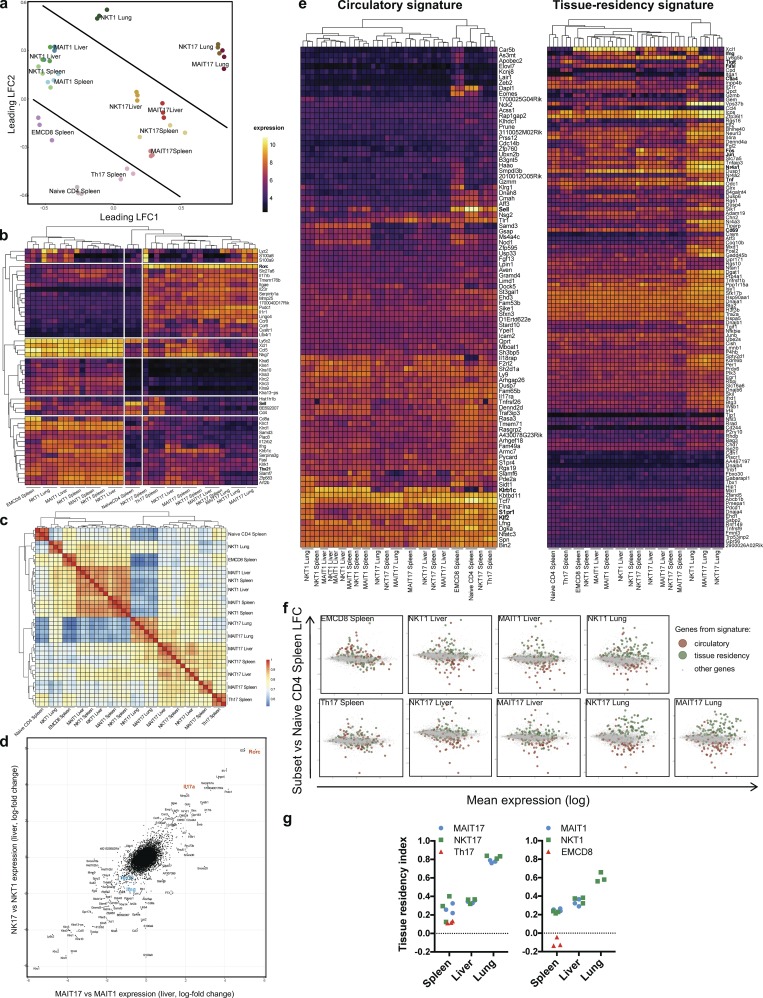
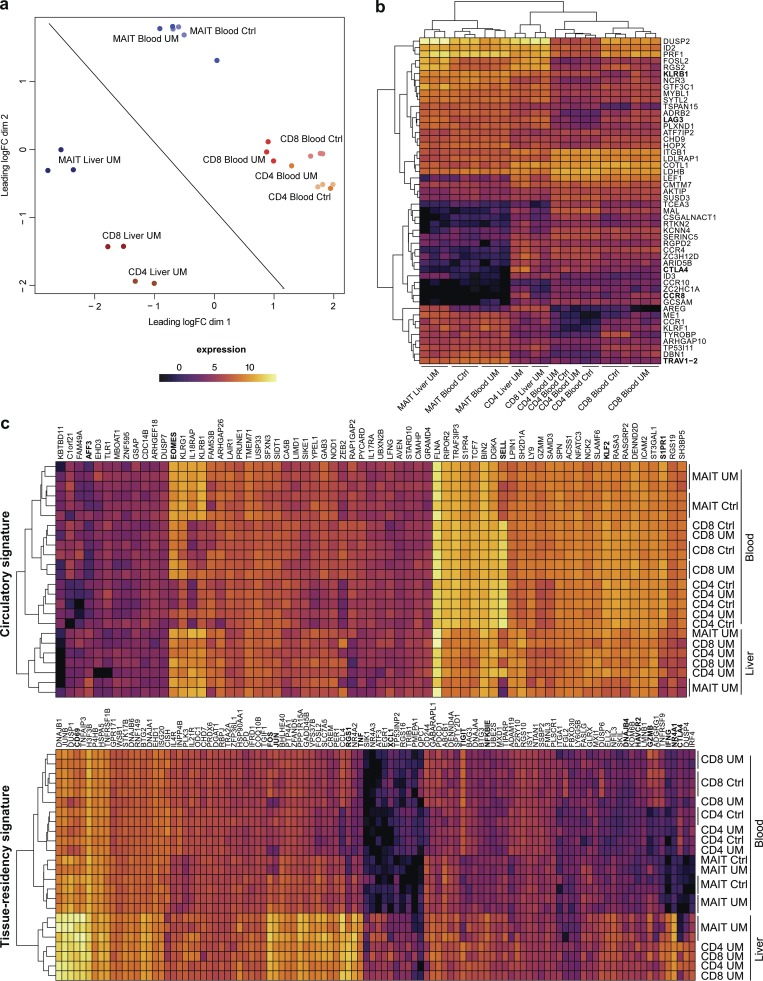
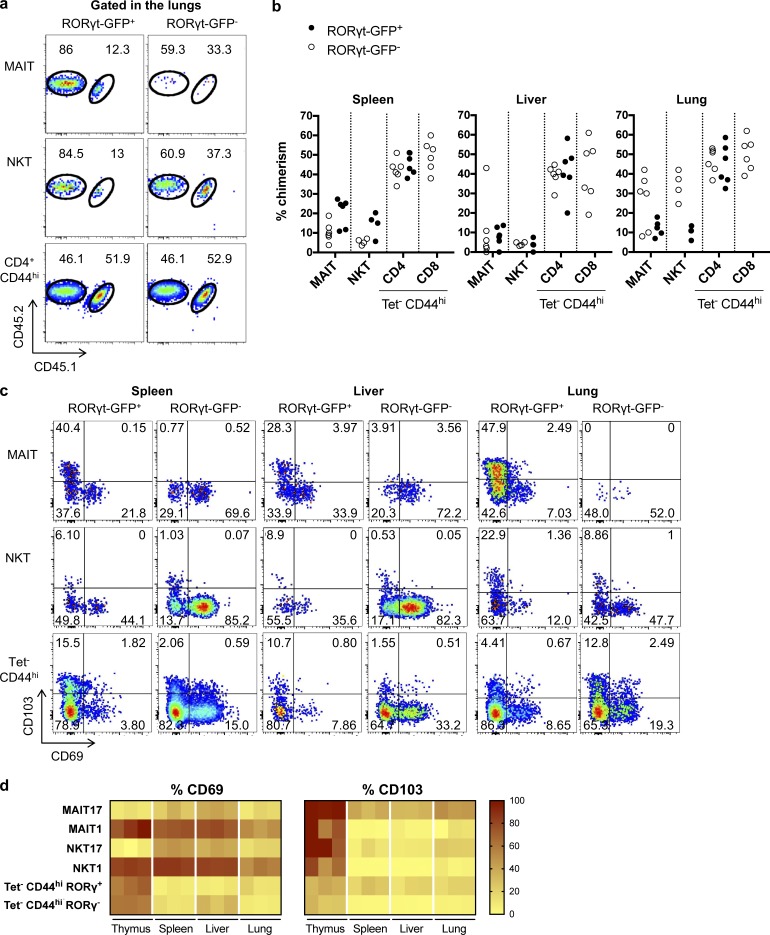
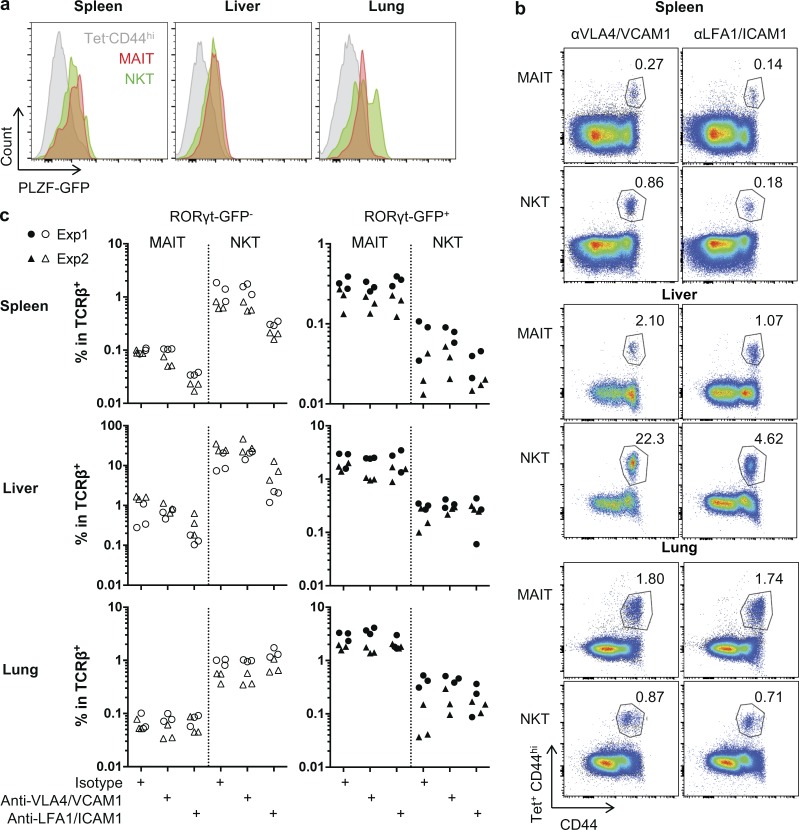
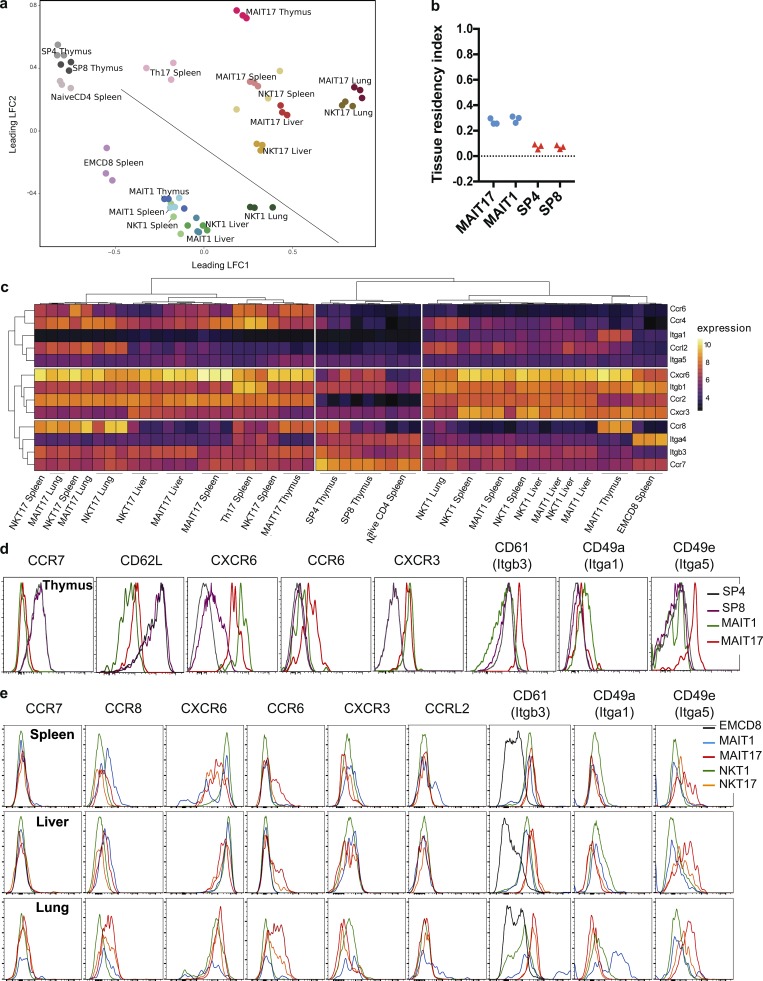
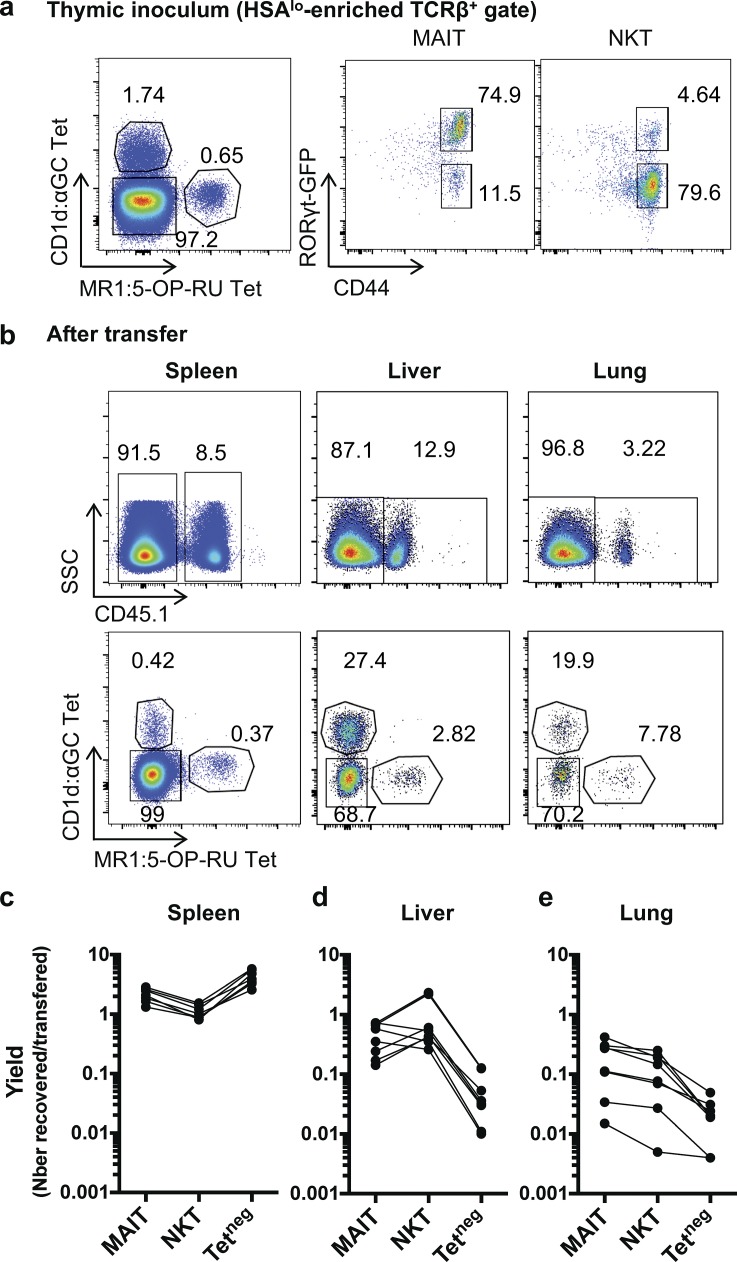
Mucosal-associated invariant T (MAIT) cells are abundant T cells with unique specificity for microbial metabolites. MAIT conservation along evolution indicates important functions, but their low frequency in mice has hampered their detailed characterization. Here, we performed the first transcriptomic analysis of murine MAIT cells. MAIT1 (RORγtneg) and MAIT17 (RORγt+) subsets were markedly distinct from mainstream T cells, but quasi-identical to NKT1 and NKT17 subsets. The expression of similar programs was further supported by strong correlations of MAIT and NKT frequencies in various organs. In both mice and humans, MAIT subsets expressed gene signatures associated with tissue residency. Accordingly, parabiosis experiments demonstrated that MAIT and NKT cells are resident in the spleen, liver, and lungs, with LFA1/ICAM1 interactions controlling MAIT1 and NKT1 retention in spleen and liver. The transcriptional program associated with tissue residency was already expressed in thymus, as confirmed by adoptive transfer experiments. Altogether, shared thymic differentiation processes generate "preset" NKT and MAIT subsets with defined effector functions, associated with specific positioning into tissues.
© 2018 Salou et al.
Figures
References
-
- Beura L.K., Mitchell J.S., Thompson E.A., Schenkel J.M., Mohammed J., Wijeyesinghe S., Fonseca R., Burbach B.J., Hickman H.D., Vezys V., et al. . 2018. Intravital mucosal imaging of CD8+ resident memory T cells shows tissue-autonomous recall responses that amplify secondary memory. Nat. Immunol. 19:173–182. 10.1038/s41590-017-0029-3 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
