

High-yield production of human Dicer by transfection of human HEK293-EBNA1 cells grown in suspension
- PMID: 30522464
- PMCID: PMC6282390
- DOI: 10.1186/s12896-018-0485-3
High-yield production of human Dicer by transfection of human HEK293-EBNA1 cells grown in suspension
Abstract
Background: Dicer is a 219-kDa protein that plays key roles in gene regulation, particularly as the ribonuclease III enzyme responsible for cleaving precursor miRNA substrates. Its enzymatic activity is highly regulated by protein factors, and this regulation can impact on the levels of miRNAs and modulate the behavior of a cell. To better understand the underlying mechanisms of regulation, detailed enzymatic and structural characterization of Dicer are needed. However, these types of studies generally require several milligrams of recombinant protein, and efficient preparation of such quantities of pure human Dicer remains a challenge. To prepare large quantities of human Dicer, we have optimized transfection in HEK293-6E cells grown in suspension and streamlined a purification procedure.
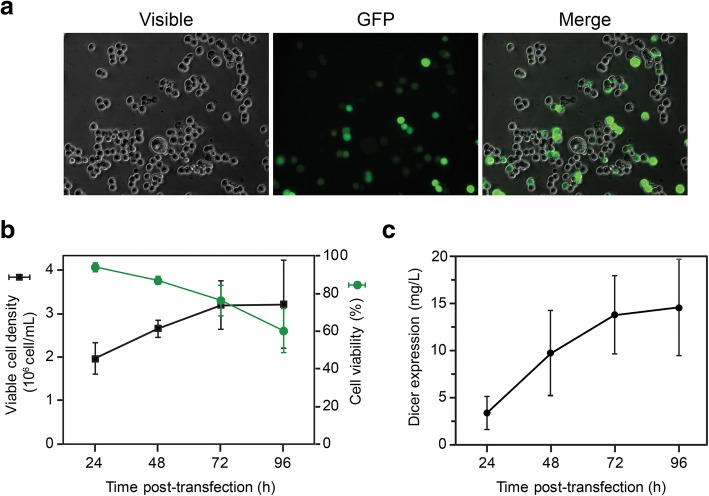
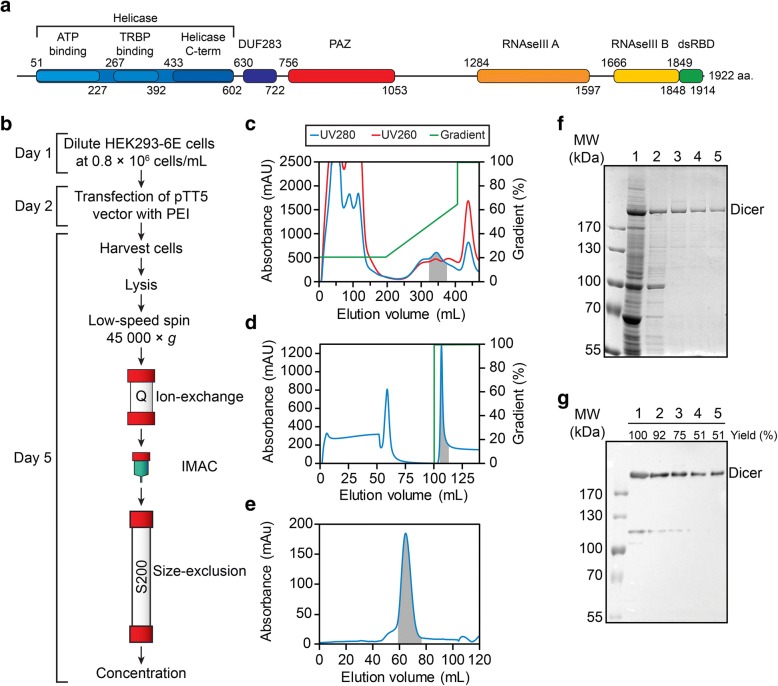
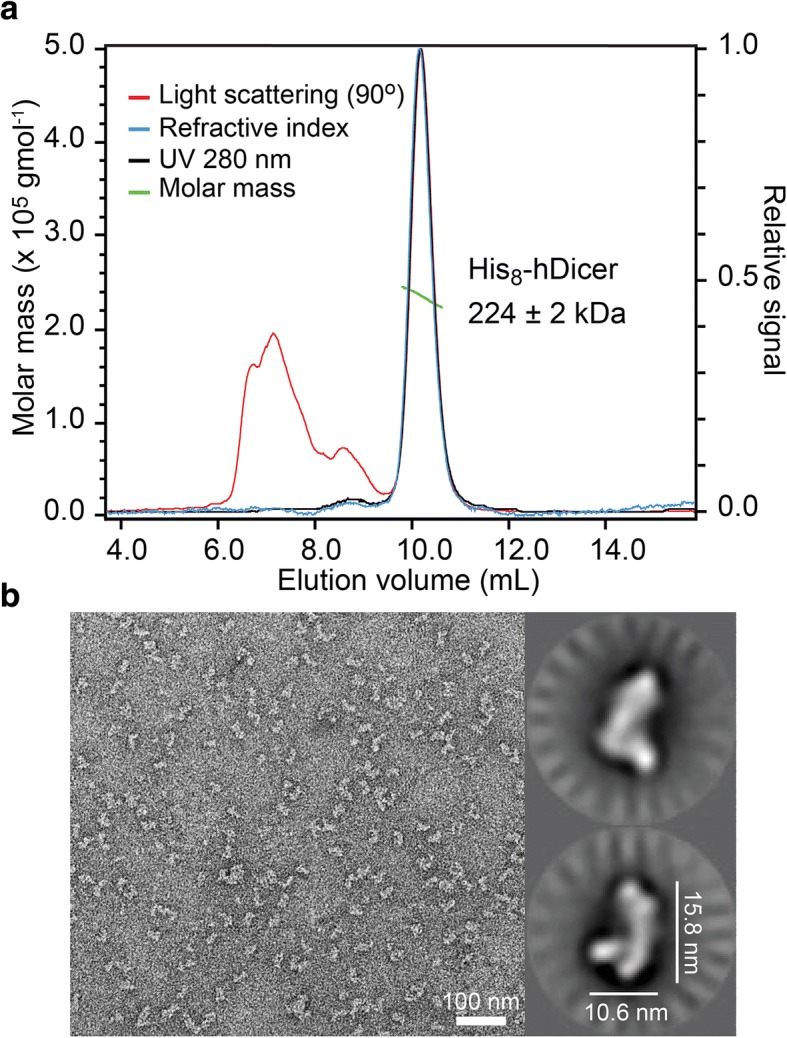
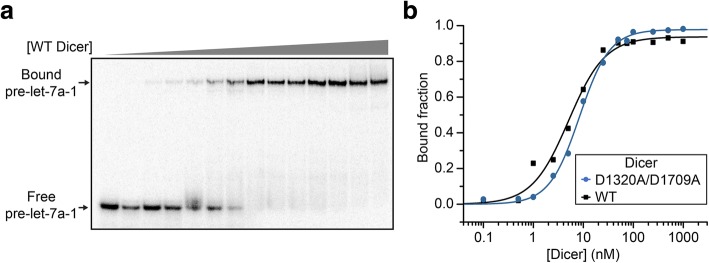
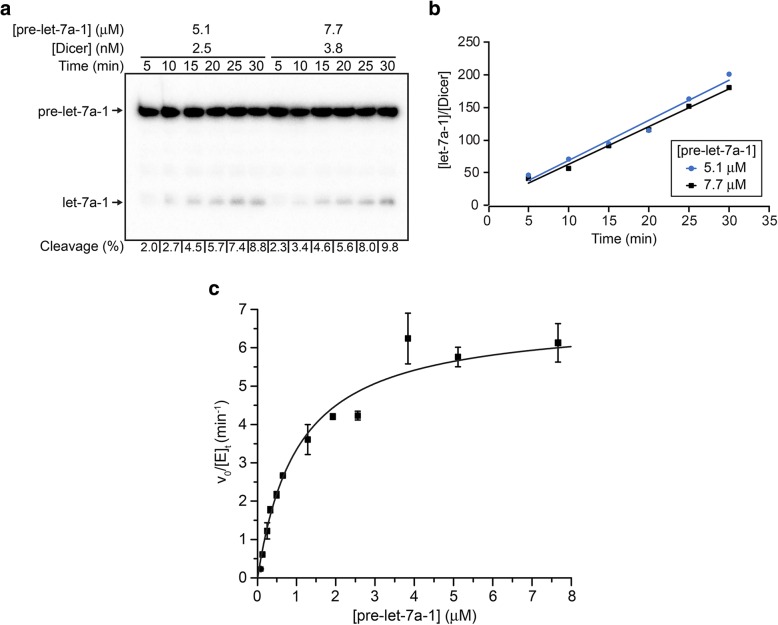
Results: Transfection conditions were first optimized to achieve expression levels between 10 and 18 mg of recombinant Dicer per liter of culture. A three-step purification protocol was then developed that yields 4-9 mg of purified Dicer per liter of culture in a single day. From SEC-MALS/RI analysis and negative stain TEM, we confirmed that the purified protein is monomerically pure ( ≥ 98%) and folds with the characteristic L-shape geometry. Using an electrophoretic mobility shift assay, a dissociation constant (Kd) of 5 nM was measured for Dicer binding to pre-let-7a-1, in agreement with previous reports. However, when probing the cleavage activity of Dicer for pre-let-7a-1, we measured kcat (7.2 ± 0.5 min- 1) and KM (1.2 ± 0.3 μM) values that are much higher than previously reported due to experimental conditions that better respect the steady-state assumption.
Conclusions: The expression and purification protocols described here provide high yields of monomerically pure and active human Dicer. Cleavage studies of a pre-let-7 substrate with this purified Dicer reveal higher kcat and KM values than previously reported and support the current view that conformational changes are associated with substrate binding. Large quantities of highly pure Dicer will be valuable for future biochemical, biophysical and structural investigations of this key protein of the miRNA pathway.
Keywords: Dicer binding; Dicer cleavage assay; Dicer expression; Dicer purification; Human HEK293-EBNA1 cells; Mammalian cell suspension culture; Pre-let-7; Pre-miRNA.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Foulkes WD, Priest JR, Duchaine TF. DICER1: mutations, microRNAs and mechanisms. Nat Rev Cancer. 2014;14(10):662–672. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
