

Automated profiling of growth cone heterogeneity defines relations between morphology and motility
- PMID: 30523041
- PMCID: PMC6314545
- DOI: 10.1083/jcb.201711023
Automated profiling of growth cone heterogeneity defines relations between morphology and motility
Abstract
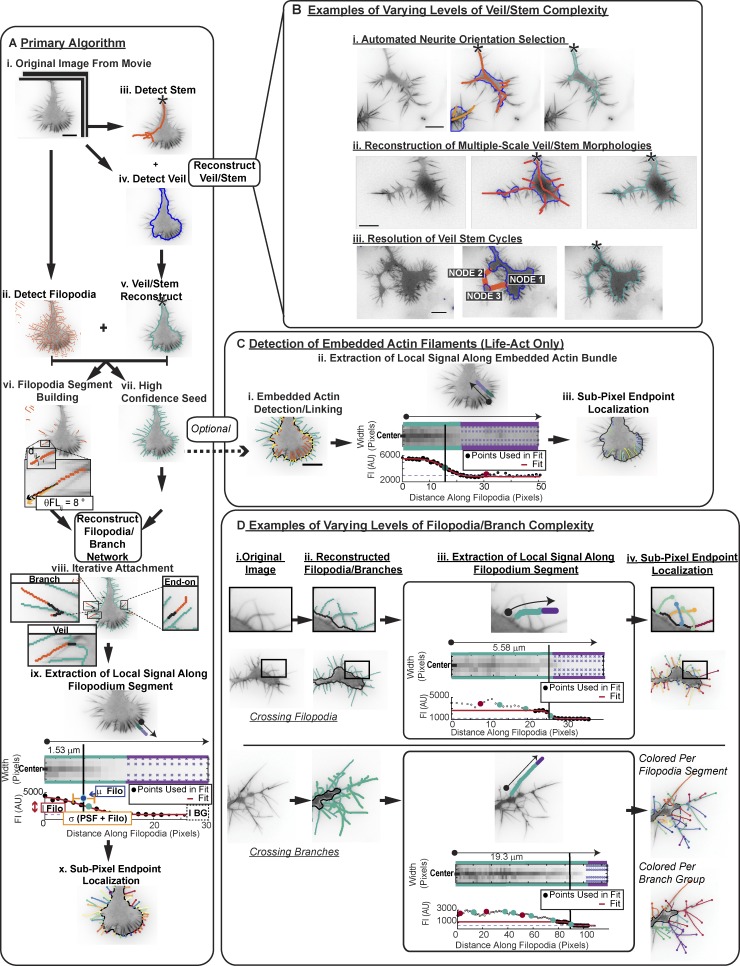
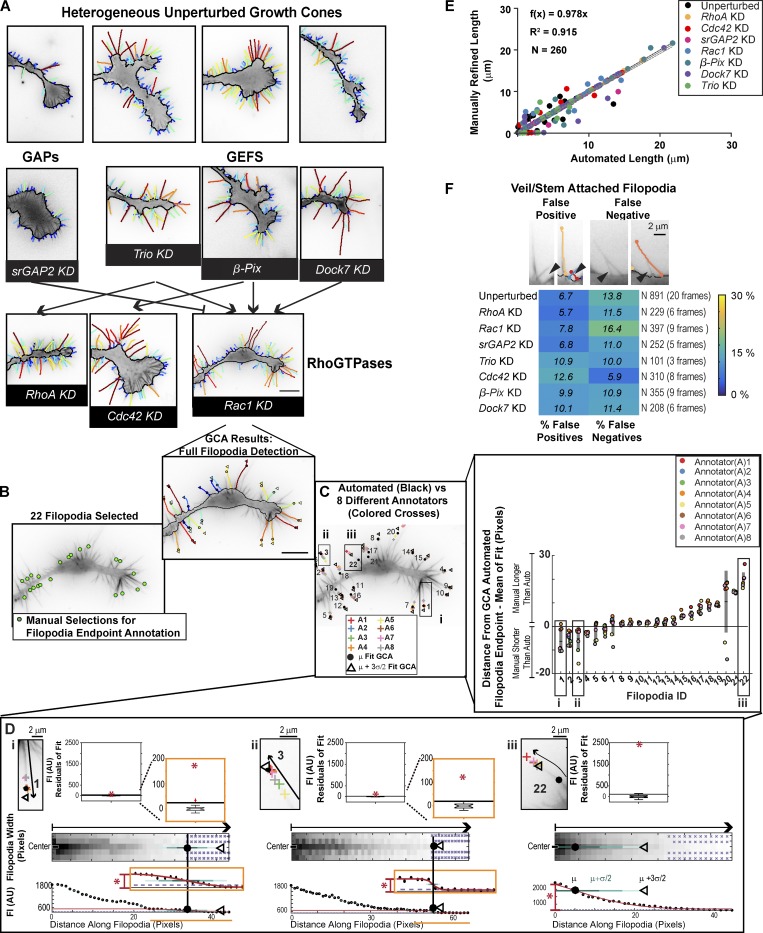
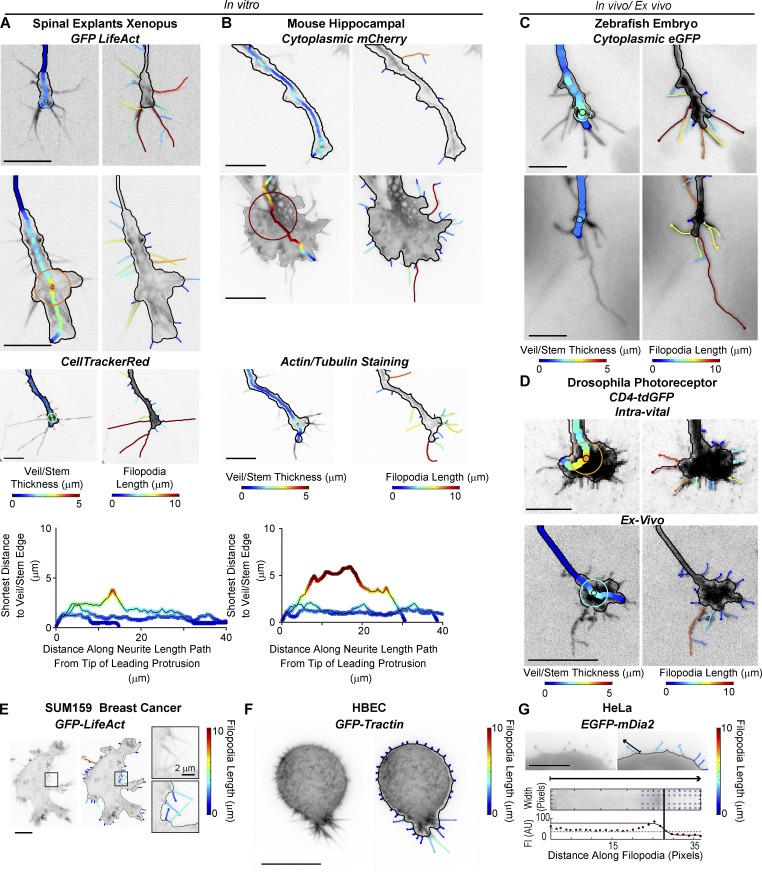
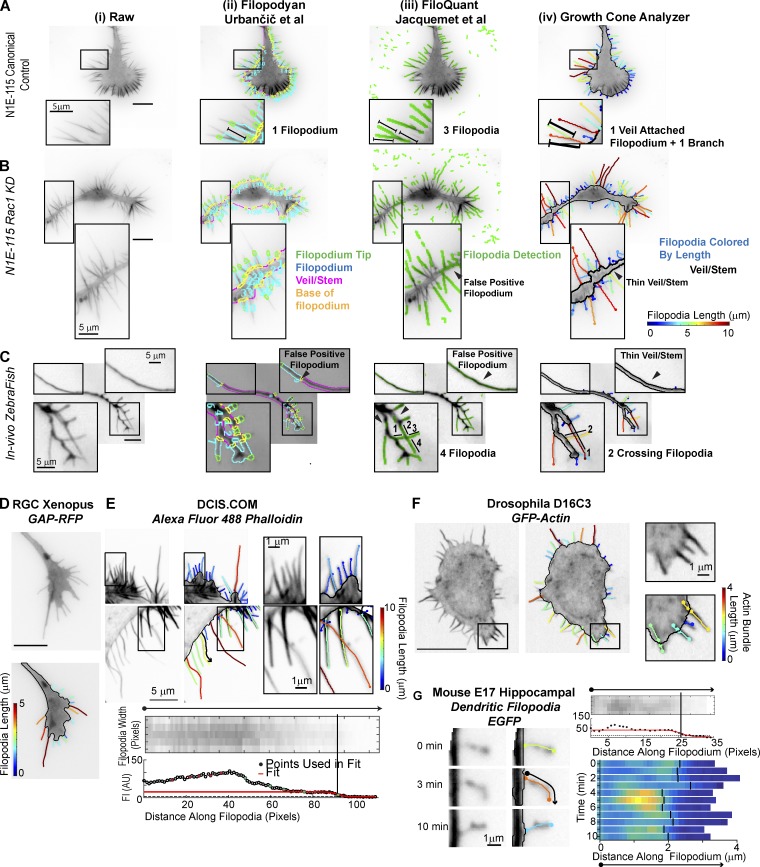
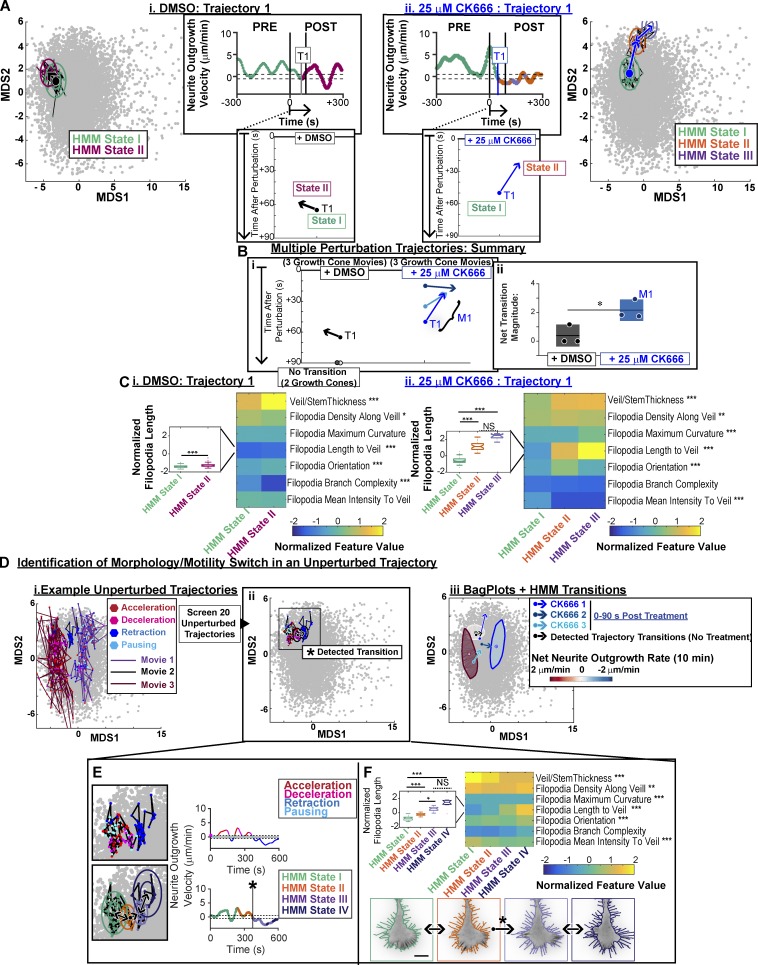
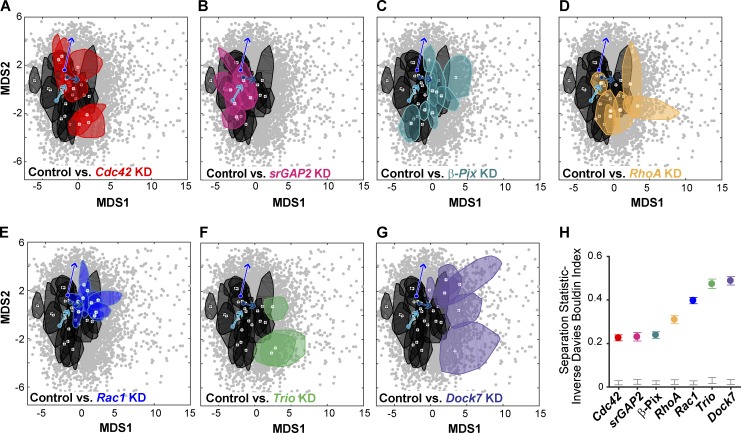
Growth cones are complex, motile structures at the tip of an outgrowing neurite. They often exhibit a high density of filopodia (thin actin bundles), which complicates the unbiased quantification of their morphologies by software. Contemporary image processing methods require extensive tuning of segmentation parameters, require significant manual curation, and are often not sufficiently adaptable to capture morphology changes associated with switches in regulatory signals. To overcome these limitations, we developed Growth Cone Analyzer (GCA). GCA is designed to quantify growth cone morphodynamics from time-lapse sequences imaged both in vitro and in vivo, but is sufficiently generic that it may be applied to nonneuronal cellular structures. We demonstrate the adaptability of GCA through the analysis of growth cone morphological variation and its relation to motility in both an unperturbed system and in the context of modified Rho GTPase signaling. We find that perturbations inducing similar changes in neurite length exhibit underappreciated phenotypic nuance at the scale of the growth cone.
© 2018 Bagonis et al.
Figures
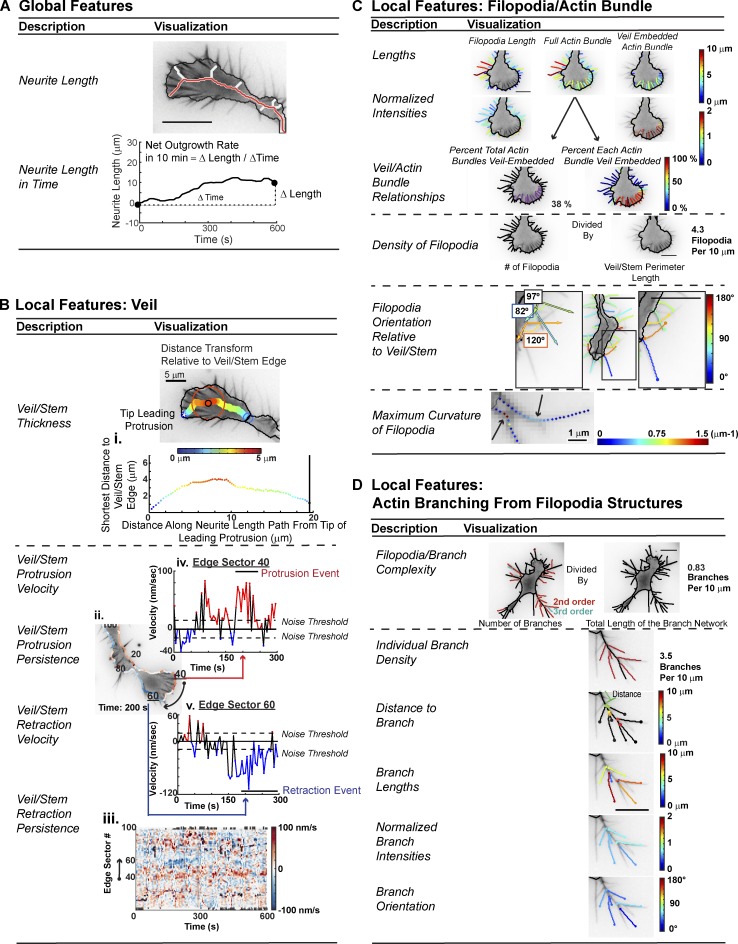
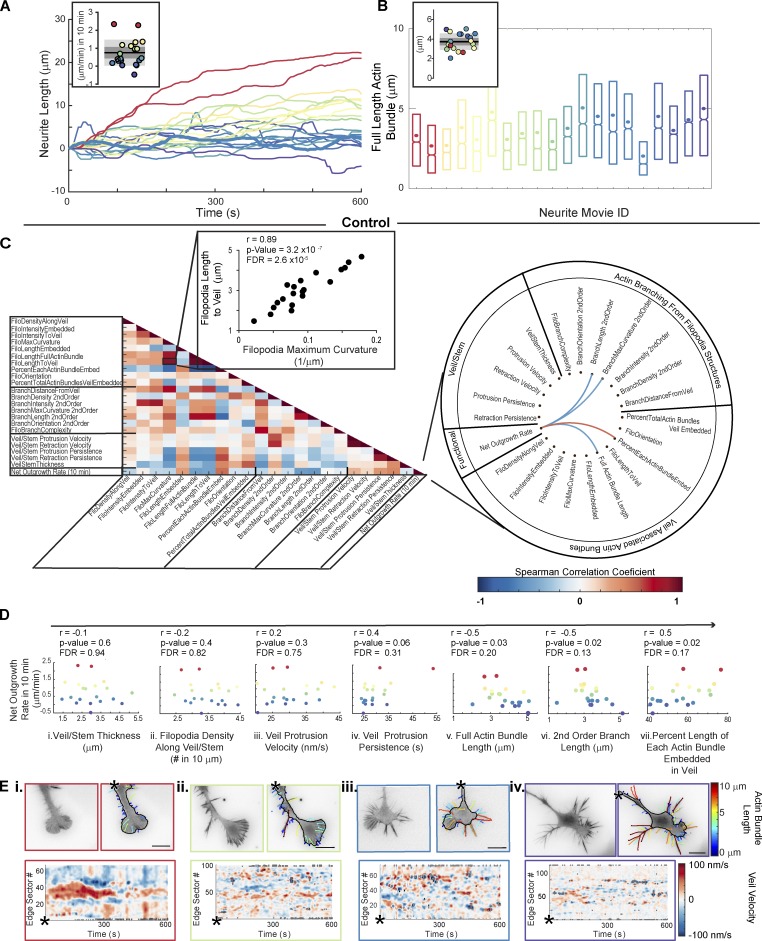
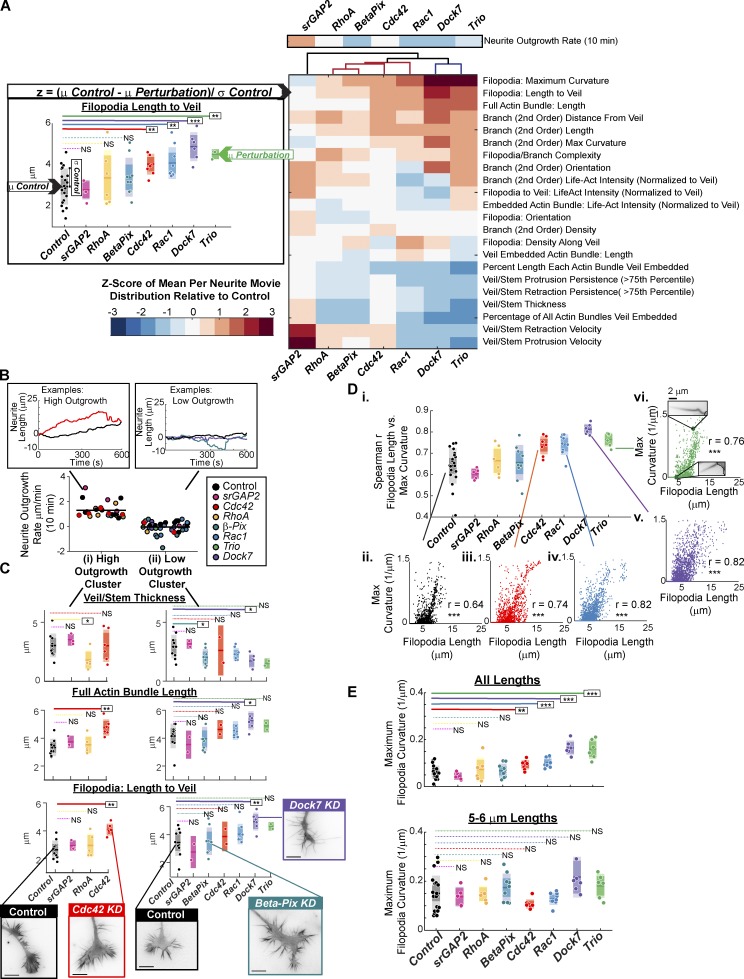
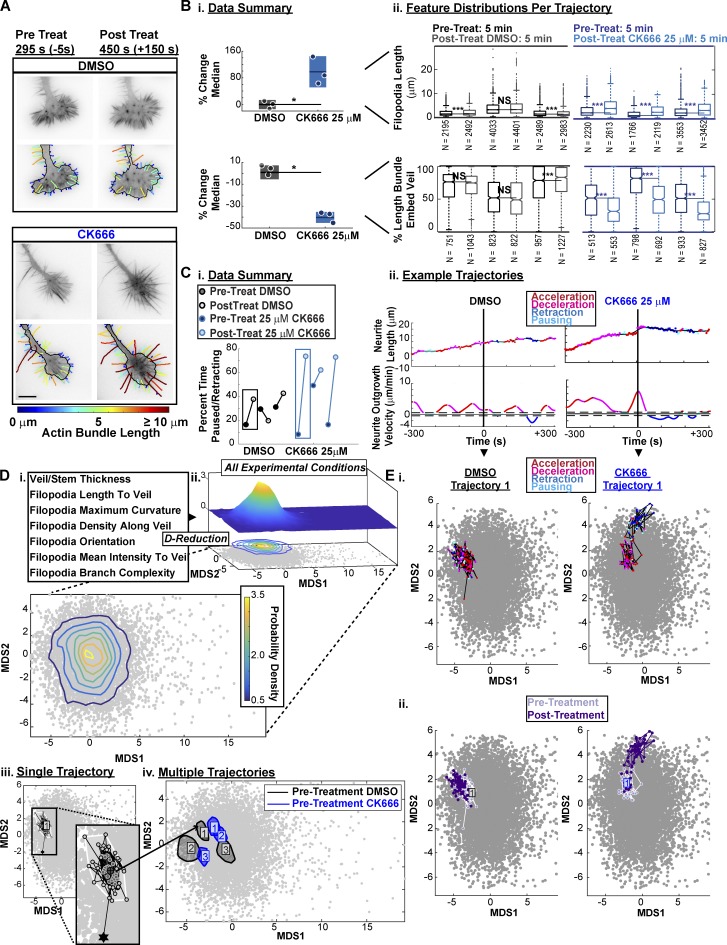
References
-
- Avis D. 1983. A survey of heuristics for the weighted matching problem. Networks. 13:475–493. 10.1002/net.3230130404 - DOI
-
- Benjamini Y., and Hochberg Y.. 1995. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Series B Stat. Methodol. 57:289–300.
-
- Borg I. 1997. Modern Multidimensional Scaling: Theory and Applications. Springer-Verlag, New York: 614 pp.
