

Commensal-specific T cell plasticity promotes rapid tissue adaptation to injury
- PMID: 30523076
- PMCID: PMC7304459
- DOI: 10.1126/science.aat6280
Commensal-specific T cell plasticity promotes rapid tissue adaptation to injury
Abstract
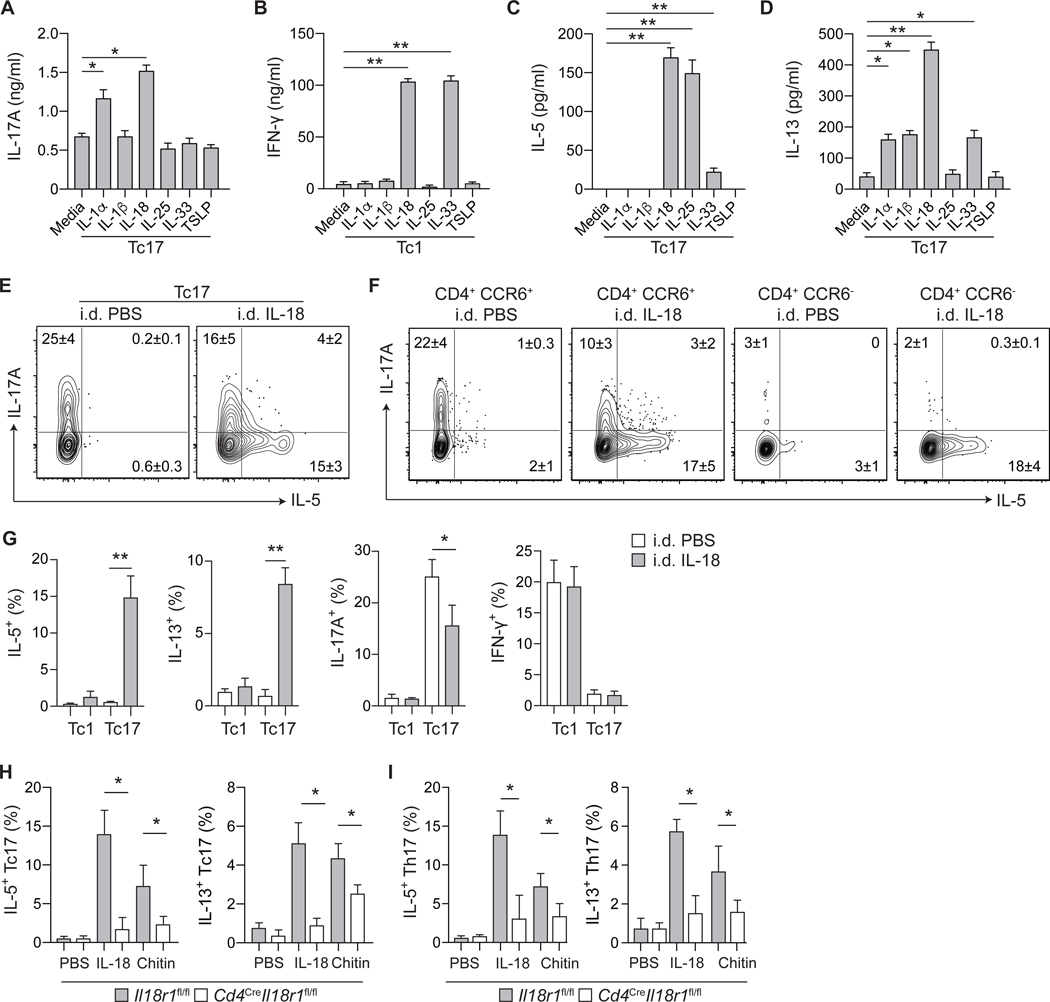
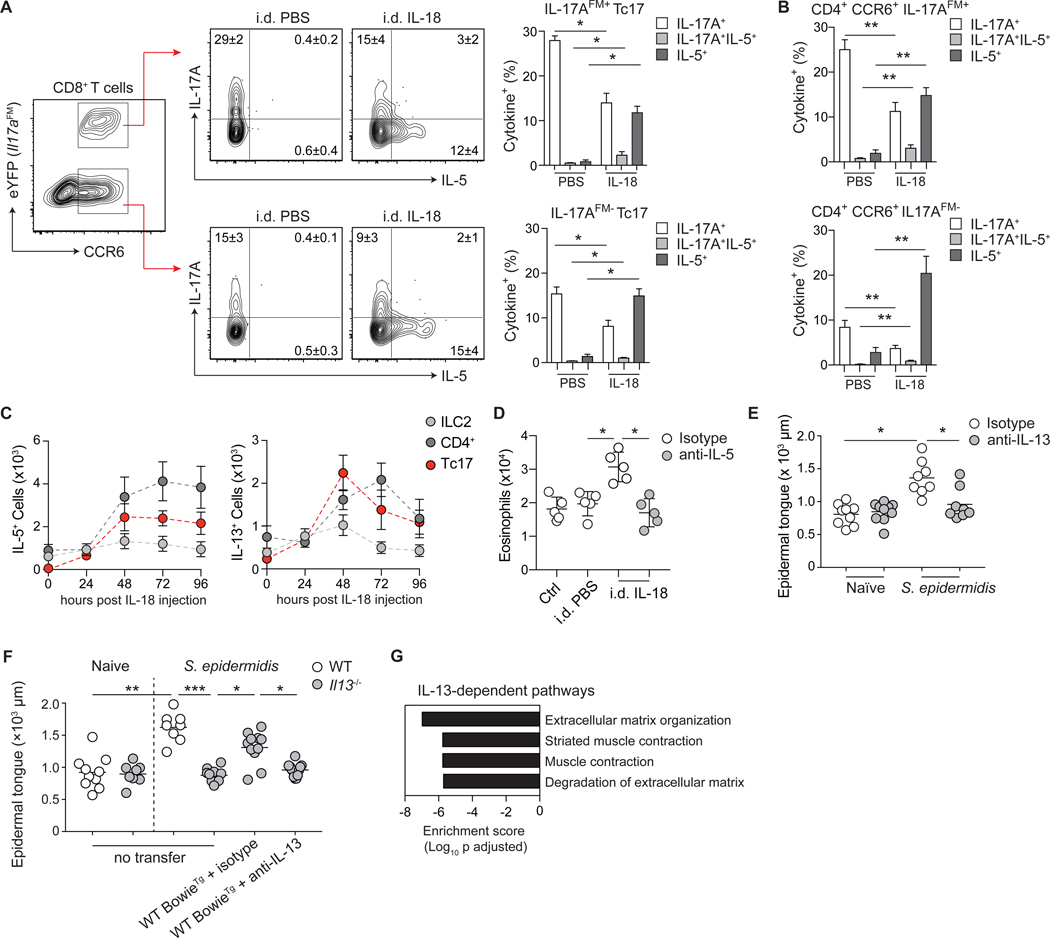
Barrier tissues are primary targets of environmental stressors and are home to the largest number of antigen-experienced lymphocytes in the body, including commensal-specific T cells. We found that skin-resident commensal-specific T cells harbor a paradoxical program characterized by a type 17 program associated with a poised type 2 state. Thus, in the context of injury and exposure to inflammatory mediators such as interleukin-18, these cells rapidly release type 2 cytokines, thereby acquiring contextual functions. Such acquisition of a type 2 effector program promotes tissue repair. Aberrant type 2 responses can also be unleashed in the context of local defects in immunoregulation. Thus, commensal-specific T cells co-opt tissue residency and cell-intrinsic flexibility as a means to promote both local immunity and tissue adaptation to injury.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
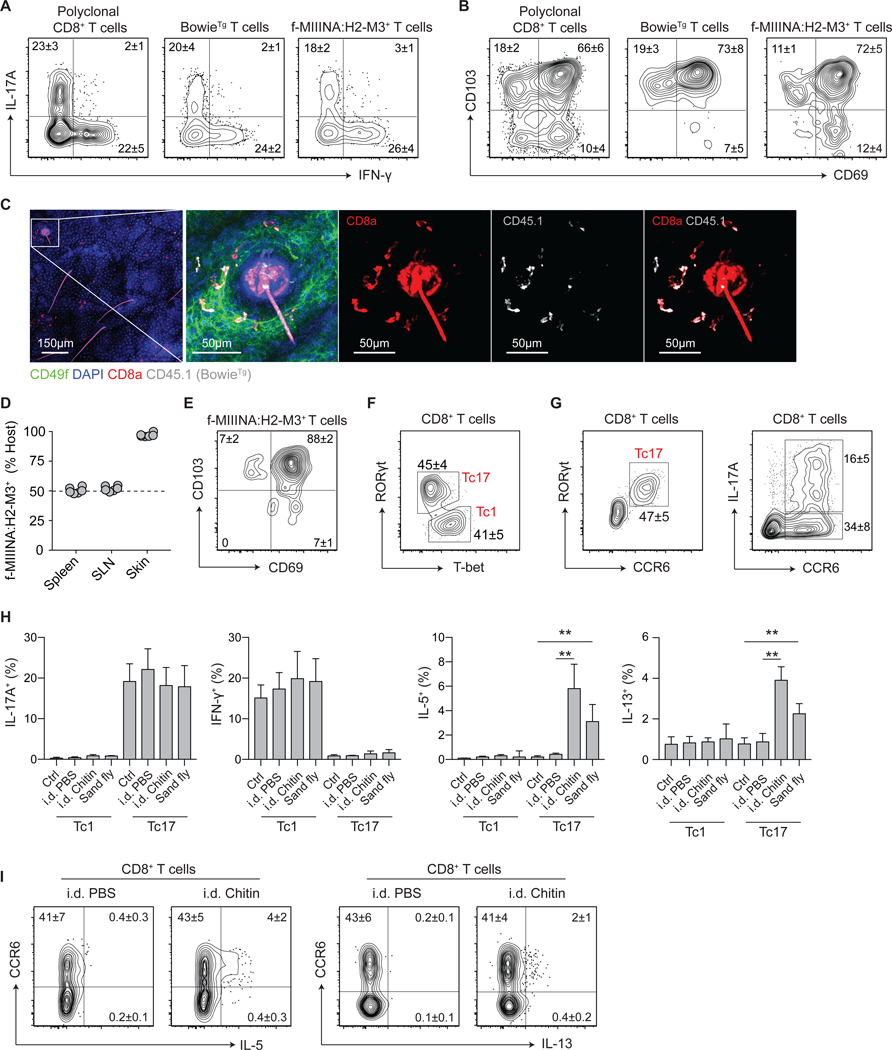
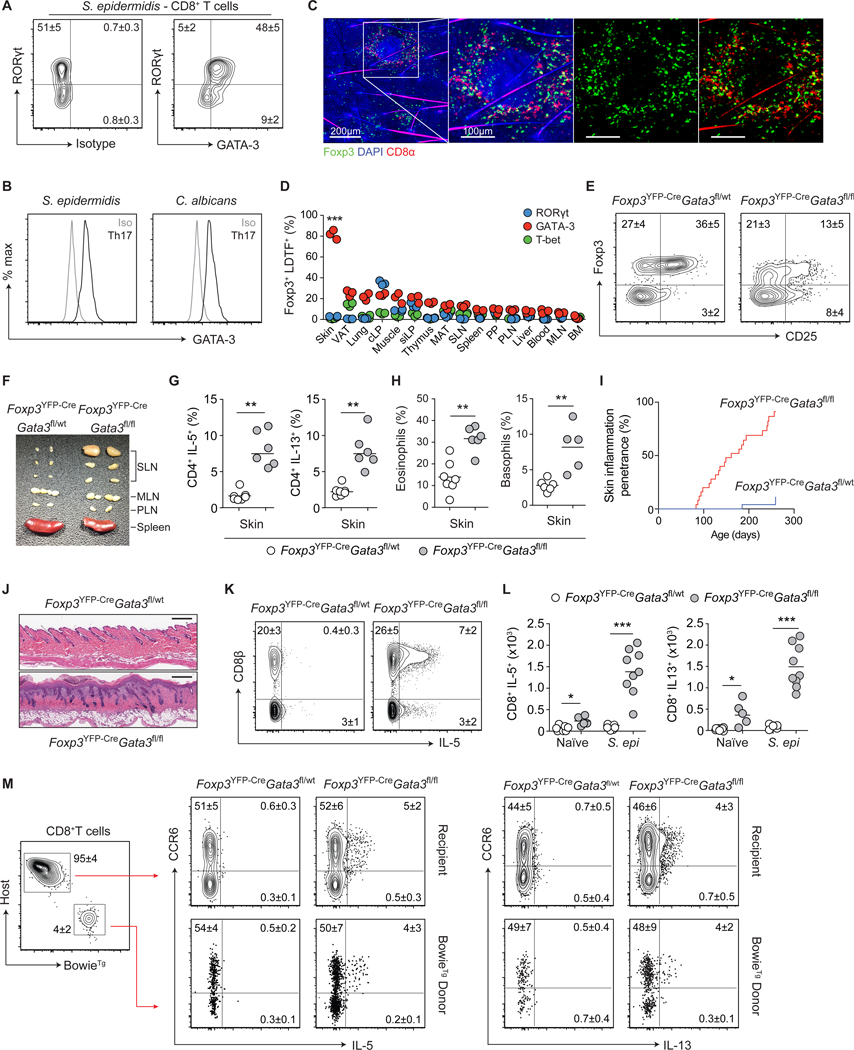
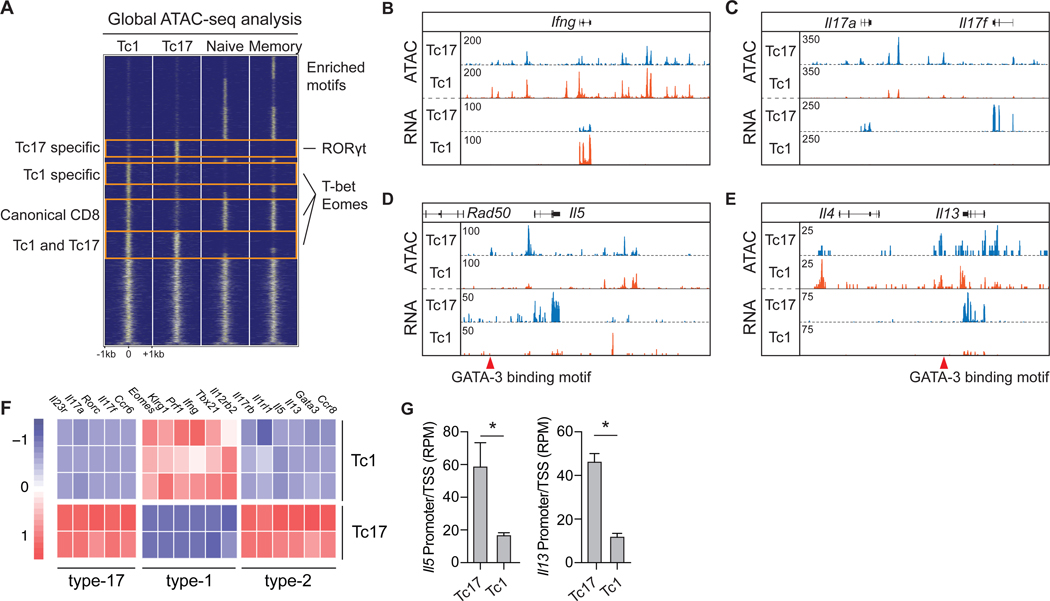
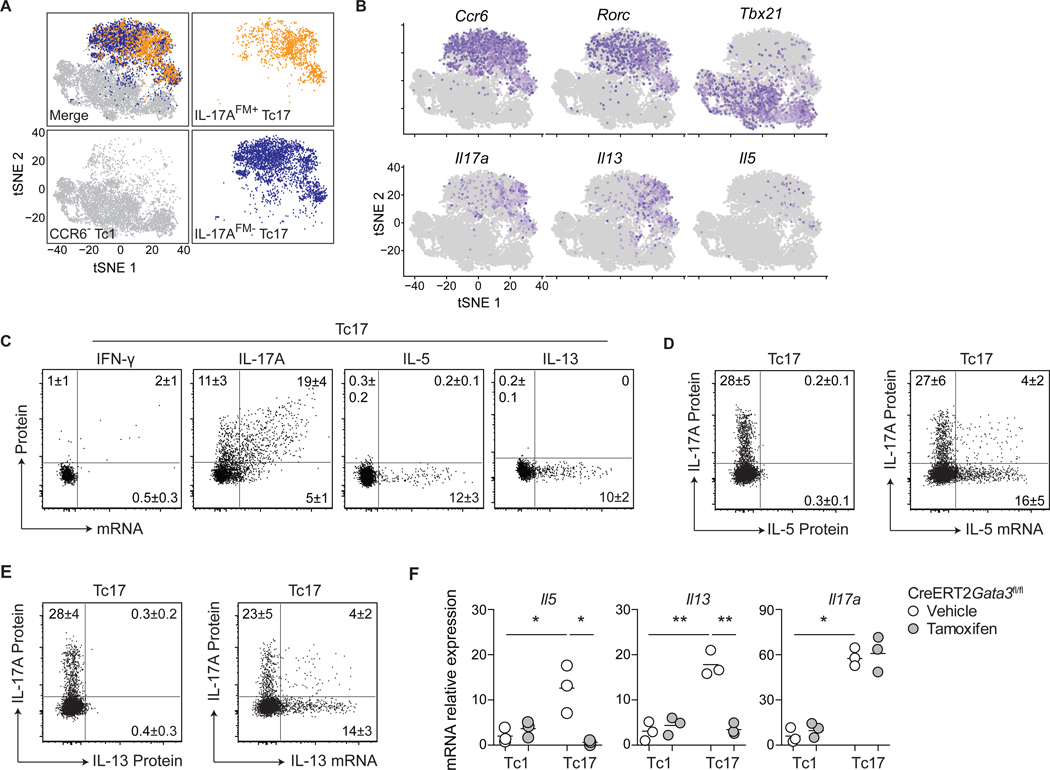
Comment in
-
Poised plasticity of skin T cells.Nat Rev Immunol. 2019 Feb;19(2):70-71. doi: 10.1038/s41577-018-0116-1. Nat Rev Immunol. 2019. PMID: 30610222 No abstract available.
-
Immunology: Skin T Cells Switch Identity to Protect and Heal.Curr Biol. 2019 Mar 18;29(6):R220-R223. doi: 10.1016/j.cub.2019.01.055. Curr Biol. 2019. PMID: 30889396
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
