

Spiking Pattern of the Mouse Developing Inner Hair Cells Is Mostly Invariant Along the Tonotopic Axis
- PMID: 30524238
- PMCID: PMC6262317
- DOI: 10.3389/fncel.2018.00407
Spiking Pattern of the Mouse Developing Inner Hair Cells Is Mostly Invariant Along the Tonotopic Axis
Abstract
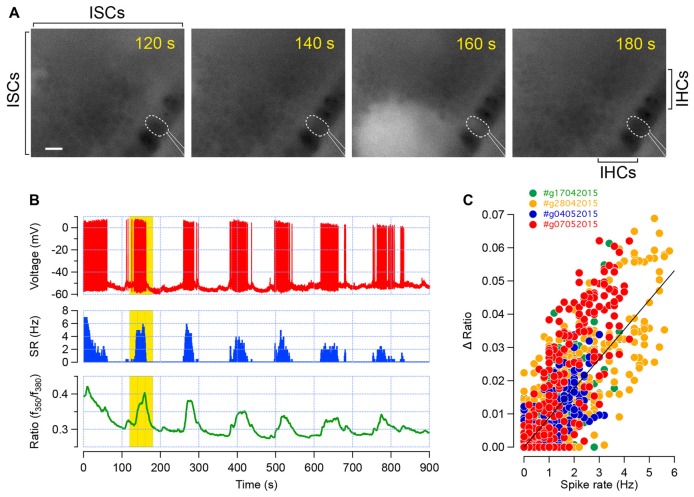
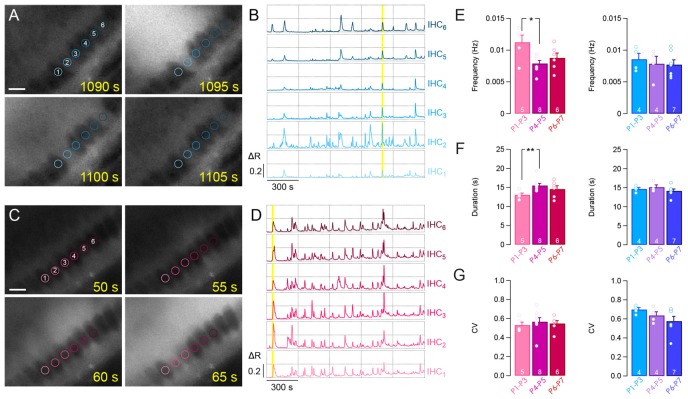
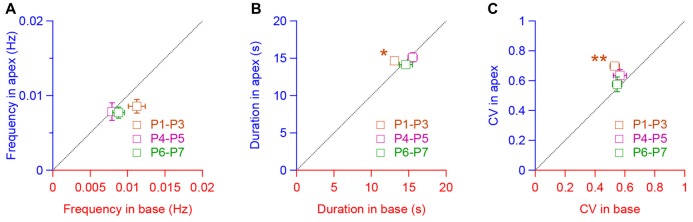
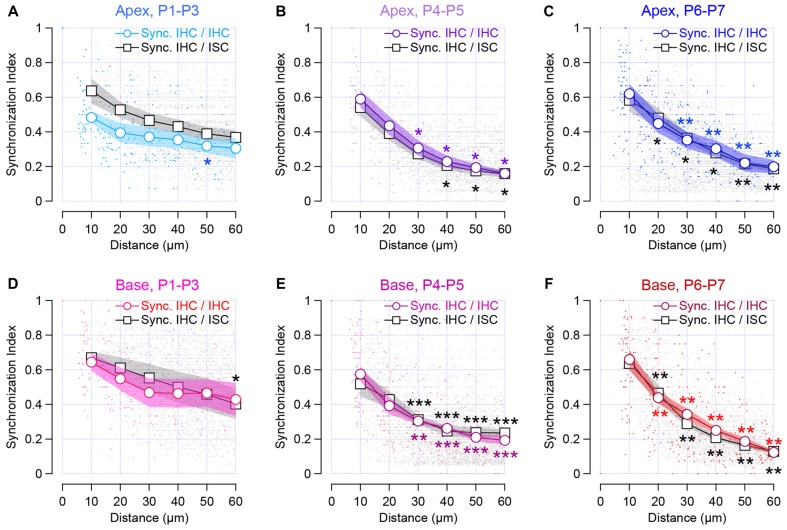
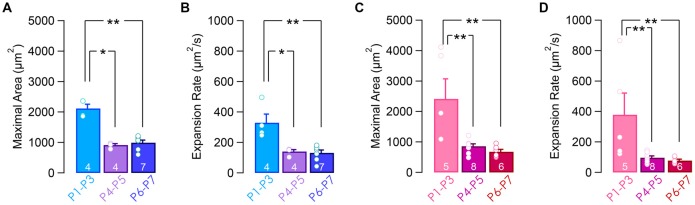
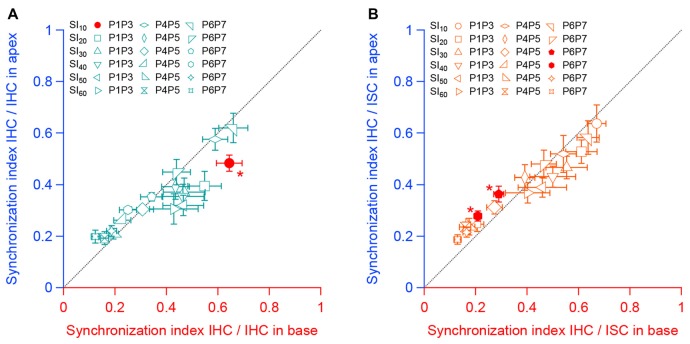
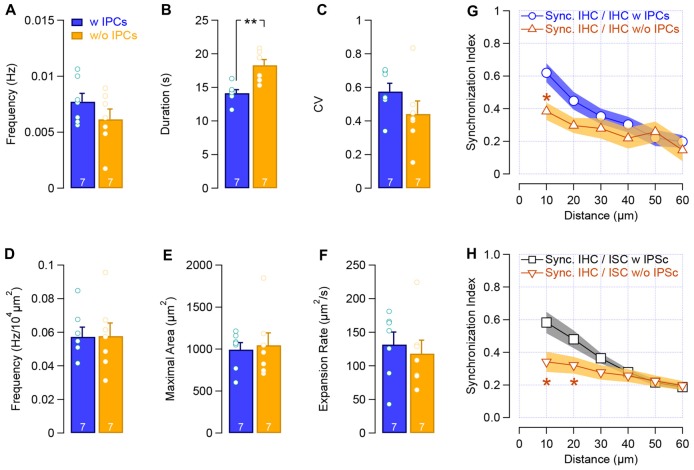
During development, the sensory cells of the cochlea, the inner hair cells (IHCs), fire spontaneous calcium action potentials. This activity at the pre-hearing stage allows the IHCs to autonomously excite the auditory nerve fibers and hence, represents an efficient mechanism to shape the tonotopic organization along the ascending auditory pathway. Using calcium imaging, we show that the activity in the developing cochlea consists of calcium waves that propagate across the supporting and sensory cells. Both basal and apical IHCs were characterized by similar spontaneous calcium transients interspaced with silent periods, consistent with bursts of action potentials recorded in patch-clamp. In addition, adjacent auditory hair cells tend to have a synchronized [Ca2+]i activity, irrespective of their location along the base-to-apex gradient of the cochlea. Finally, we show that the mechanical ablation of the inner phalangeal cells (IPCs), a class of supporting cells, reduces the synchronized [Ca2+]i activity between neighboring sensory cells. These findings support the hypothesis that the tonotopic map refinement in higher auditory centers would depend on the synchronization of a discrete number of auditory sensory cells.
Keywords: action potential; calcium transients; cochlea; sensory cells; spontaneous activity.
Figures
References
-
- Anselmi F., Hernandez V. H., Crispino G., Seydel A., Ortolano S., Roper S. D., et al. . (2008). ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc. Natl. Acad. Sci. U S A 105, 18770–18775. 10.1073/pnas.0800793105 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous