

CD137 (4-1BB) Signalosome: Complexity Is a Matter of TRAFs
- PMID: 30524423
- PMCID: PMC6262405
- DOI: 10.3389/fimmu.2018.02618
CD137 (4-1BB) Signalosome: Complexity Is a Matter of TRAFs
Abstract
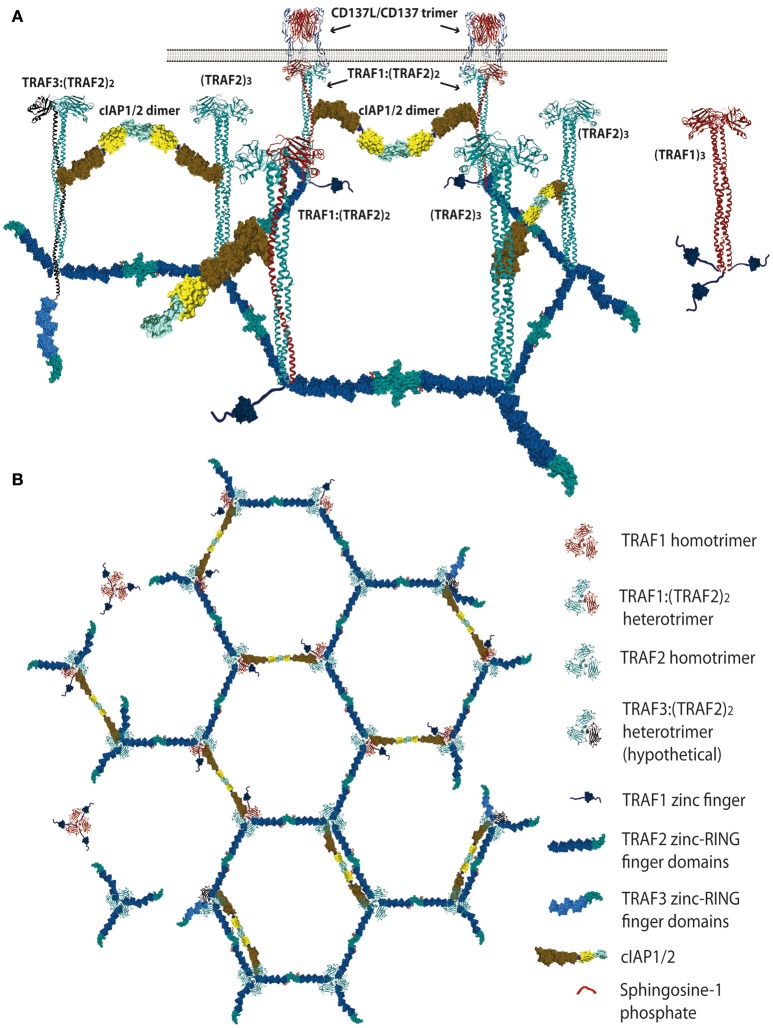
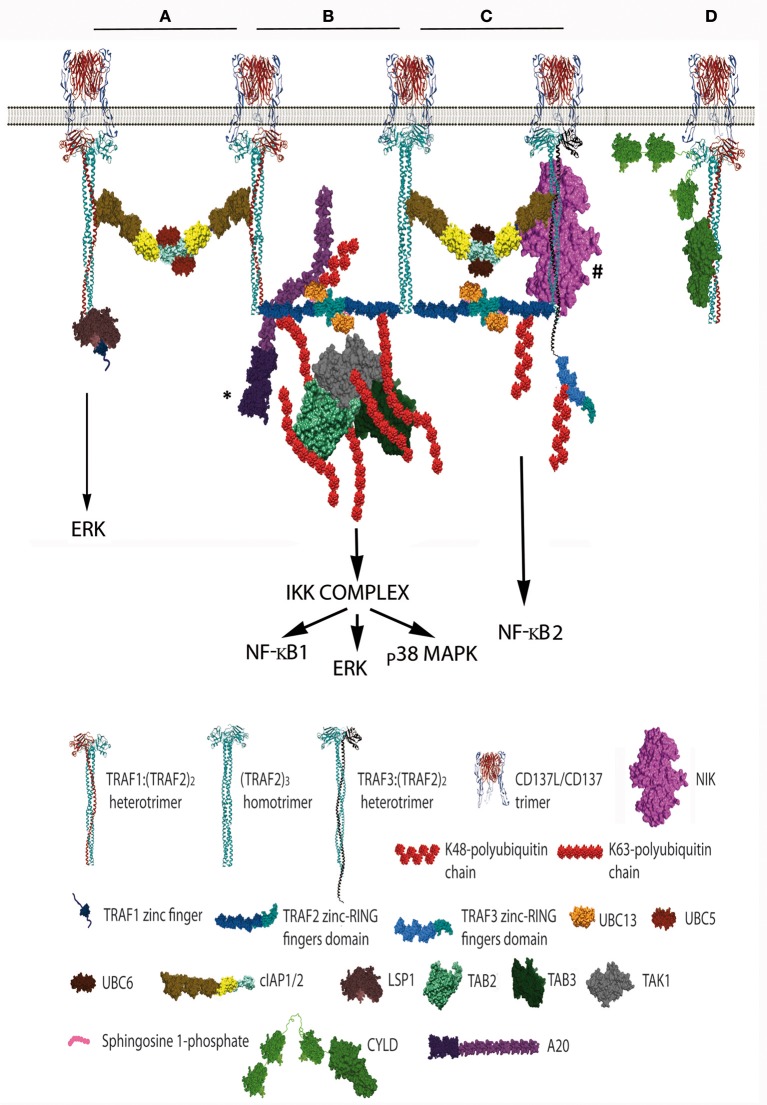
CD137 (4-1BB, Tnsfr9) is a member of the TNF-receptor (TNFR) superfamily without known intrinsic enzymatic activity in its cytoplasmic domain. Hence, akin to other members of the TNFR family, it relies on the TNFR-Associated-Factor (TRAF) family of adaptor proteins to build the CD137 signalosome for transducing signals into the cell. Thus, upon CD137 activation by binding of CD137L trimers or by crosslinking with agonist monoclonal antibodies, TRAF1, TRAF2, and TRAF3 are readily recruited to the cytoplasmic domain of CD137, likely as homo- and/or heterotrimers with different configurations, initiating the construction of the CD137 signalosome. The formation of TRAF2-RING dimers between TRAF2 molecules from contiguous trimers would help to establish a multimeric structure of TRAF-trimers that is probably essential for CD137 signaling. In addition, available studies have identified a large number of proteins that are recruited to CD137:TRAF complexes including ubiquitin ligases and proteases, kinases, and modulatory proteins. Working in a coordinated fashion, these CD137-signalosomes will ultimately promote CD137-mediated T cell proliferation and survival and will endow T cells with stronger effector functions. Current evidence allows to envision the molecular events that might take place in the early stages of CD137-signalosome formation, underscoring the key roles of TRAFs and of K63 and K48-ubiquitination of target proteins in the signaling process. Understanding the composition and fine regulation of CD137-signalosomes assembly and disassembly will be key to improve the therapeutic activities of chimeric antigen receptors (CARs) encompassing the CD137 cytoplasmic domain and a new generation of CD137 agonists for the treatment of cancer.
Keywords: 4-1BB; CD137; Immunotherapy; TNFR; TRAF1; TRAF2; TRAF3; cytotoxic T lymphocytes (CTL).
Figures
References
-
- Chung JY, Park YC, Ye H, Wu H. All TRAFs are not created equal: common and distinct molecular mechanisms of TRAF-mediated signal transduction. J Cell Sci. (2002) 115:679–88. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials