

An Experimental Pipeline for Initial Characterization of Bacterial Type III Secretion System Inhibitor Mode of Action Using Enteropathogenic Yersinia
- PMID: 30524970
- PMCID: PMC6262202
- DOI: 10.3389/fcimb.2018.00404
An Experimental Pipeline for Initial Characterization of Bacterial Type III Secretion System Inhibitor Mode of Action Using Enteropathogenic Yersinia
Abstract
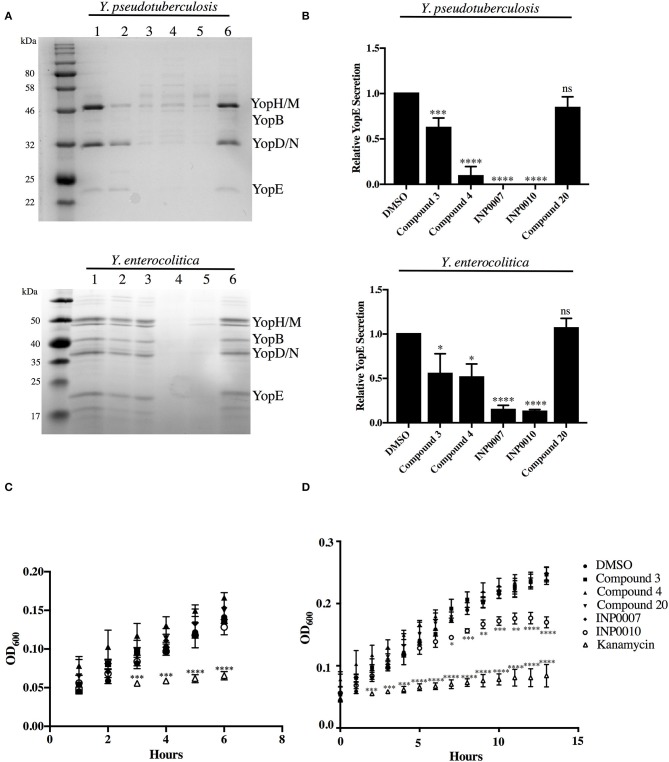
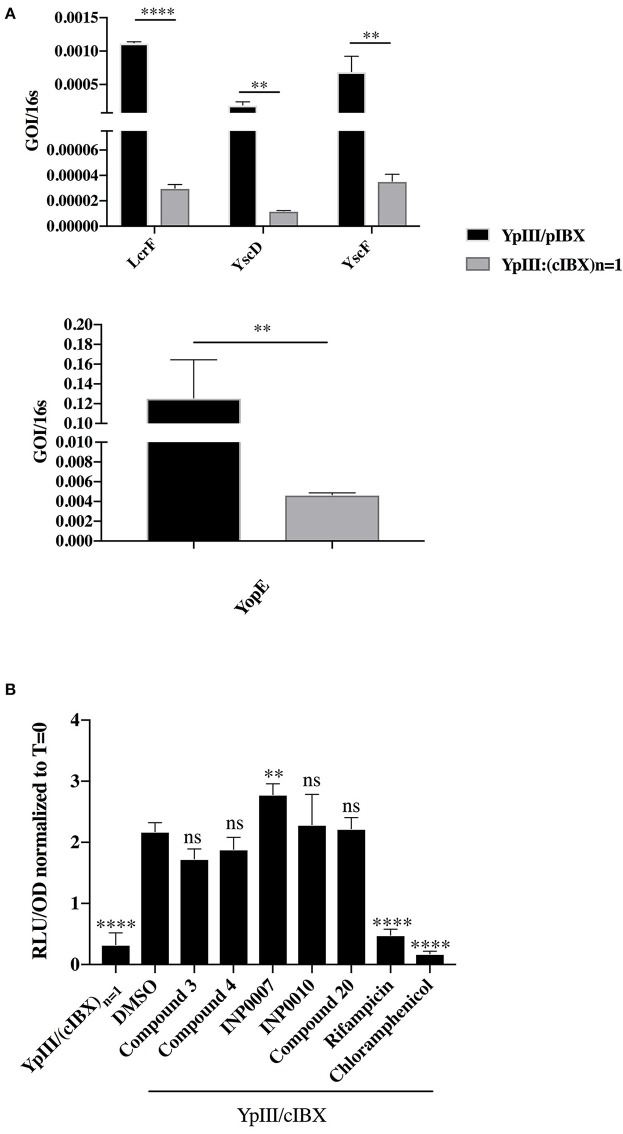
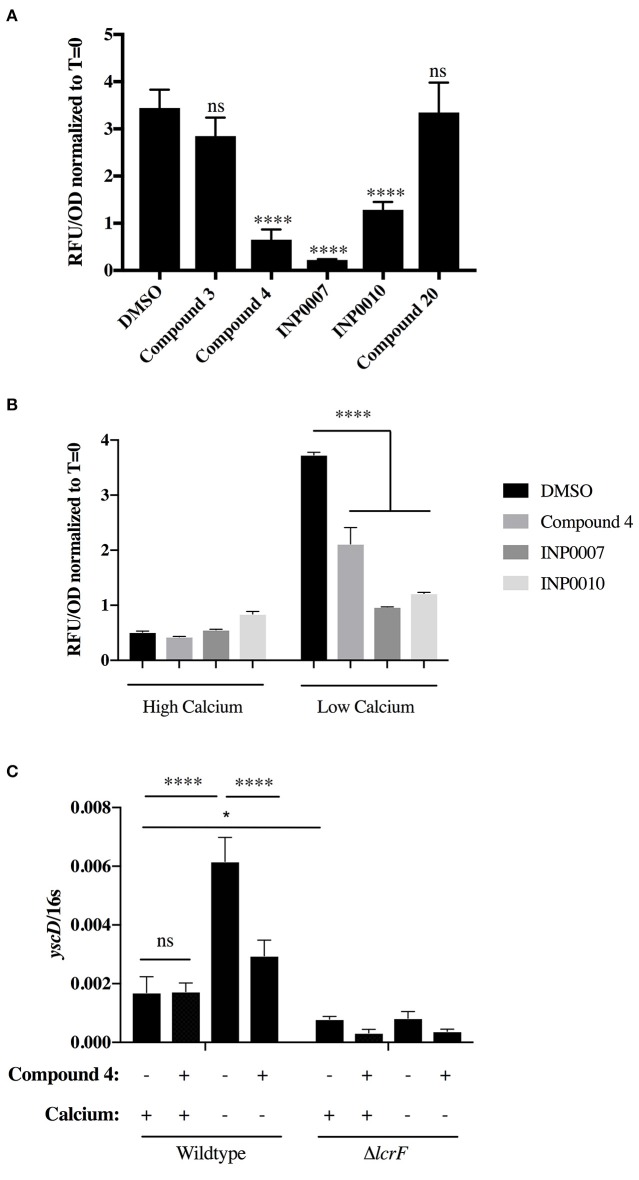
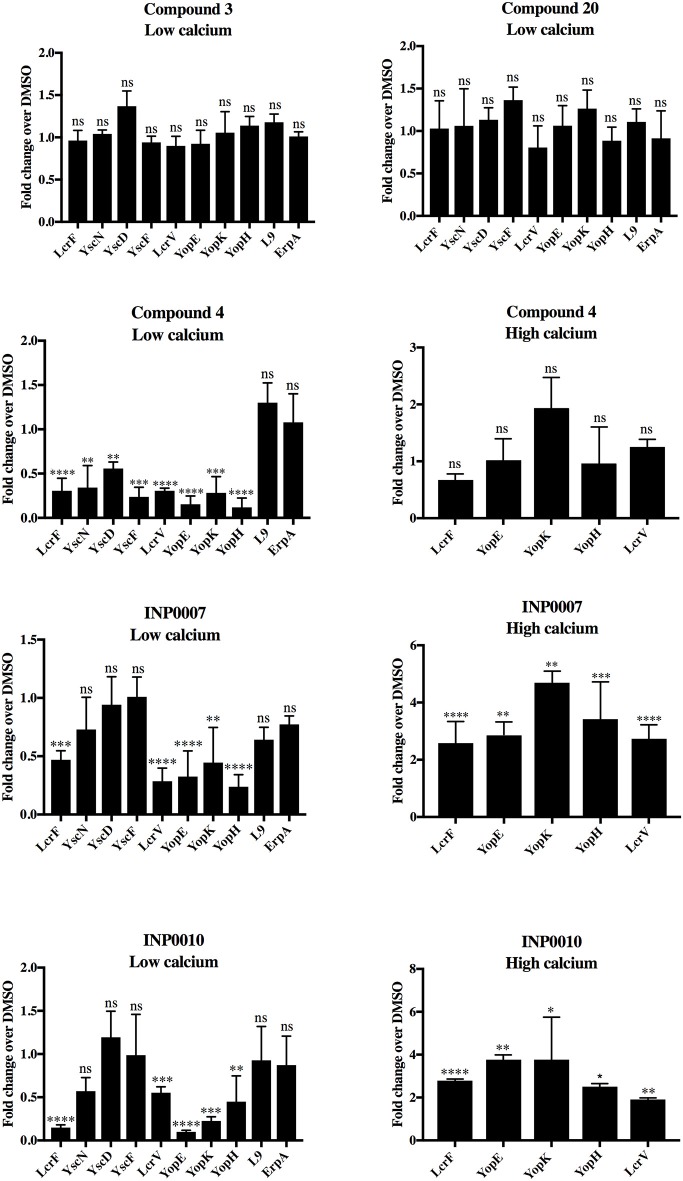
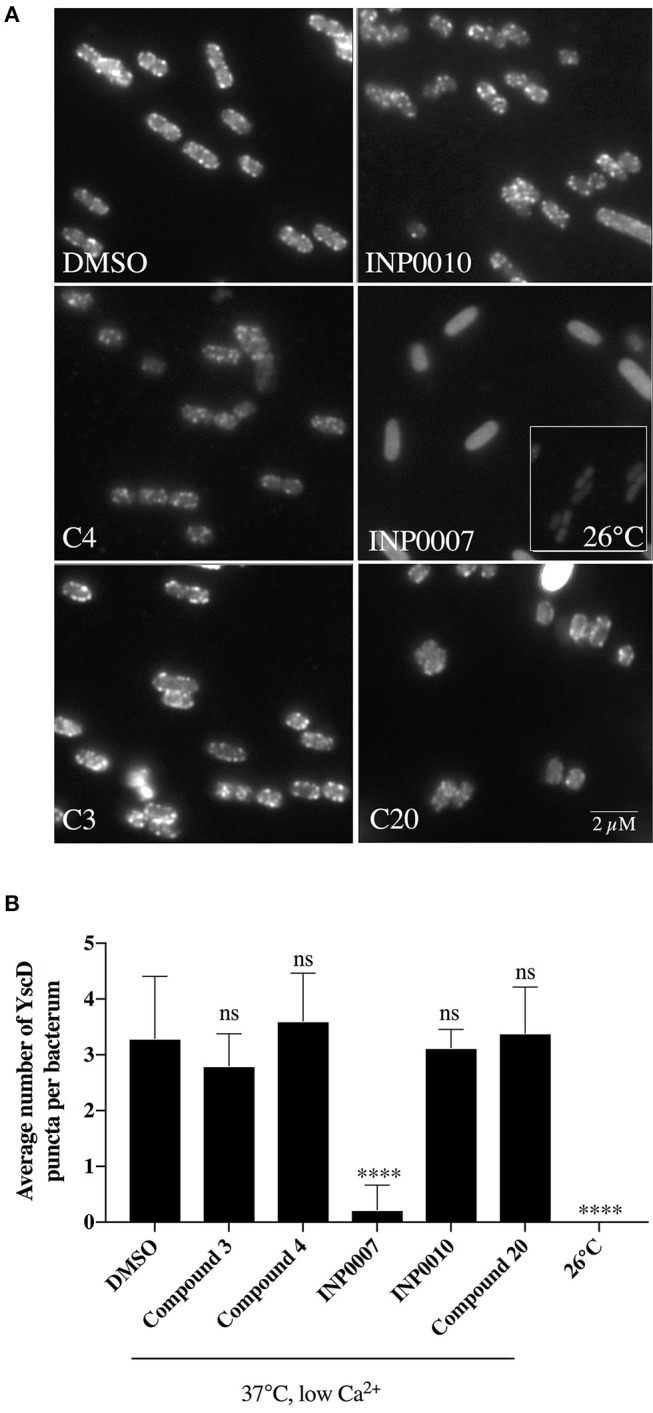
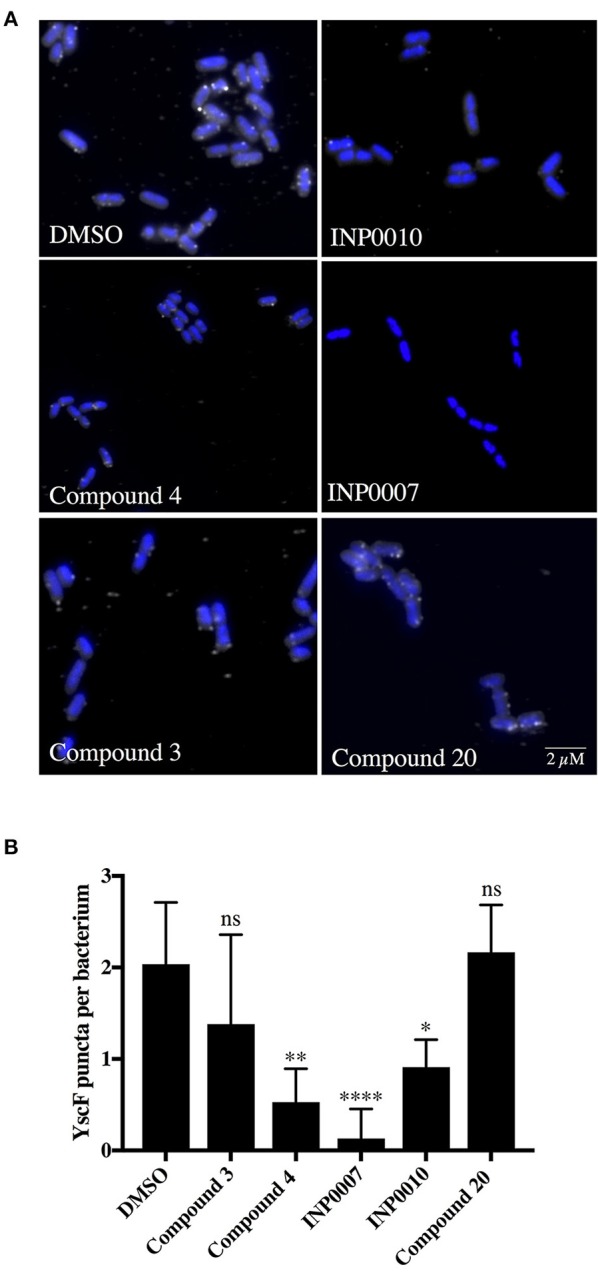
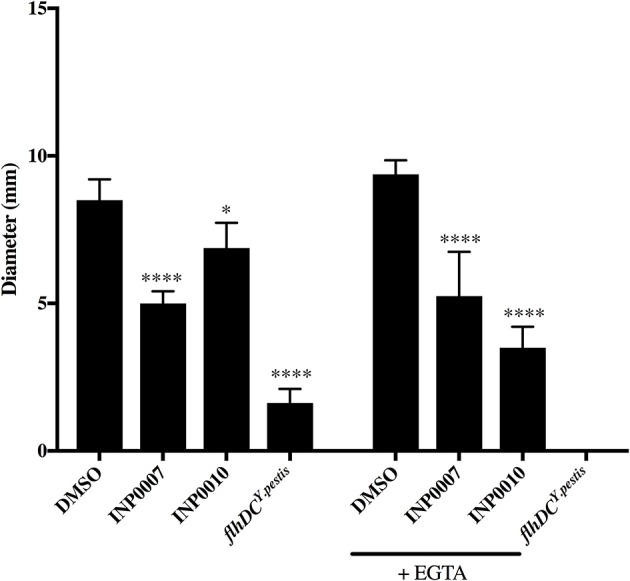
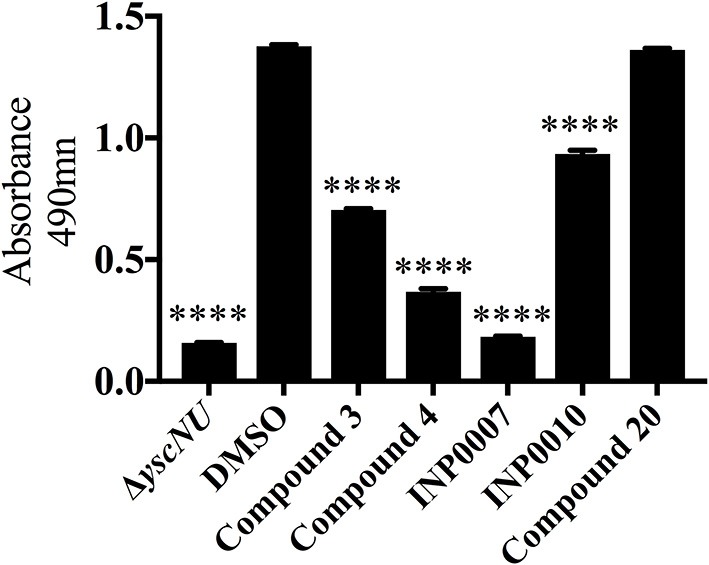
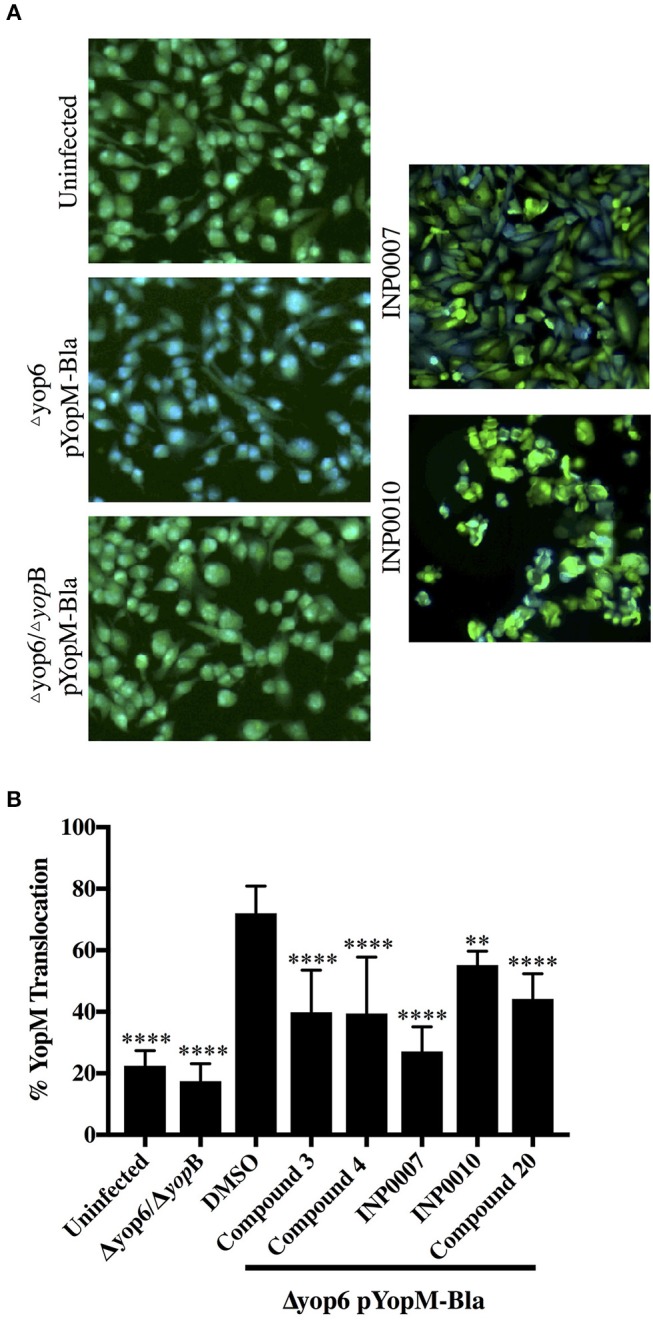
Dozens of Gram negative pathogens use one or more type III secretion systems (T3SS) to disarm host defenses or occupy a beneficial niche during infection of a host organism. While the T3SS represents an attractive drug target and dozens of compounds with T3SS inhibitory activity have been identified, few T3SS inhibitors have been validated and mode of action determined. One issue is the lack of standardized orthogonal assays following high throughput screening. Using a training set of commercially available compounds previously shown to possess T3SS inhibitory activity, we demonstrate the utility of an experiment pipeline comprised of six distinct assays to assess the stages of type III secretion impacted: T3SS gene copy number, T3SS gene expression, T3SS basal body and needle assembly, secretion of cargo through the T3SS, and translocation of T3SS effector proteins into host cells. We used enteropathogenic Yersinia as the workhorse T3SS-expressing model organisms for this experimental pipeline, as Yersinia is sensitive to all T3SS inhibitors we tested, including those active against other T3SS-expressing pathogens. We find that this experimental pipeline is capable of rapidly distinguishing between T3SS inhibitors that interrupt the process of type III secretion at different points in T3SS assembly and function. For example, our data suggests that Compound 3, a malic diamide, blocks either activity of the assembled T3SS or alters the structure of the T3SS in a way that blocks T3SS cargo secretion but not antibody recognition of the T3SS needle. In contrast, our data predicts that Compound 4, a haloid-containing sulfonamidobenzamide, disrupts T3SS needle subunit secretion or assembly. Furthermore, we suggest that misregulation of copy number control of the pYV virulence plasmid, which encodes the Yersinia T3SS, should be considered as a possible mode of action for compounds with T3SS inhibitory activity against Yersinia.
Keywords: T3SS; T3SS inhibitor; Yersinia; pYV; type III secretion system.
Figures
References
-
- Anantharajah A., Buyck J. M., Sundin C., Tulkens P. M., Mingeot-Leclercq M. P., Van Bambeke F. (2017). Salicylidene acylhydrazides and hydroxyquinolines act as inhibitors of type three secretion systems in Pseudomonas aeruginosa by distinct mechanisms. Antimicrob. Agents Chemother. 61:e02566-16. 10.1128/AAC.02566-16 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources