

Discovery of Novel Hepatitis B Virus Nucleocapsid Assembly Inhibitors
- PMID: 30525438
- PMCID: PMC6510629
- DOI: 10.1021/acsinfecdis.8b00269
Discovery of Novel Hepatitis B Virus Nucleocapsid Assembly Inhibitors
Abstract
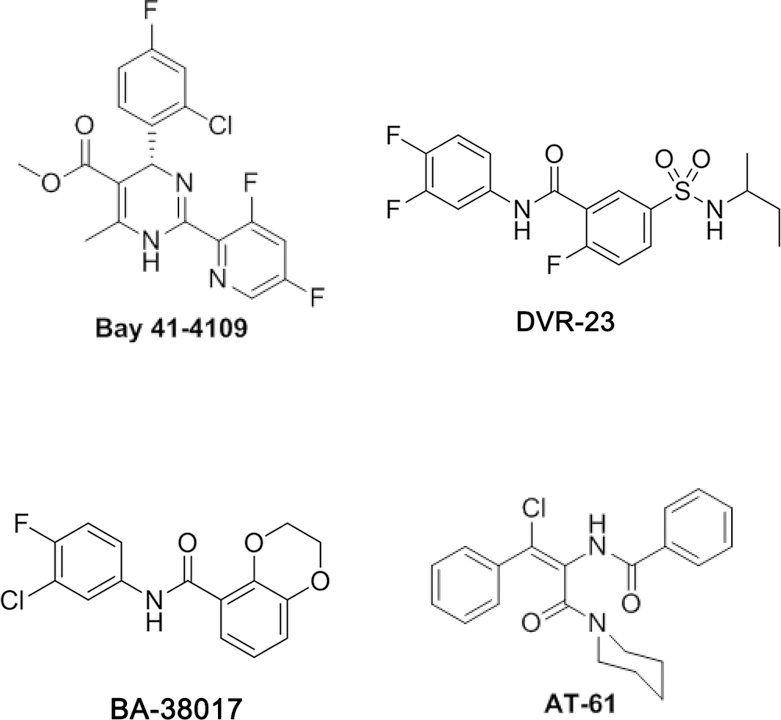
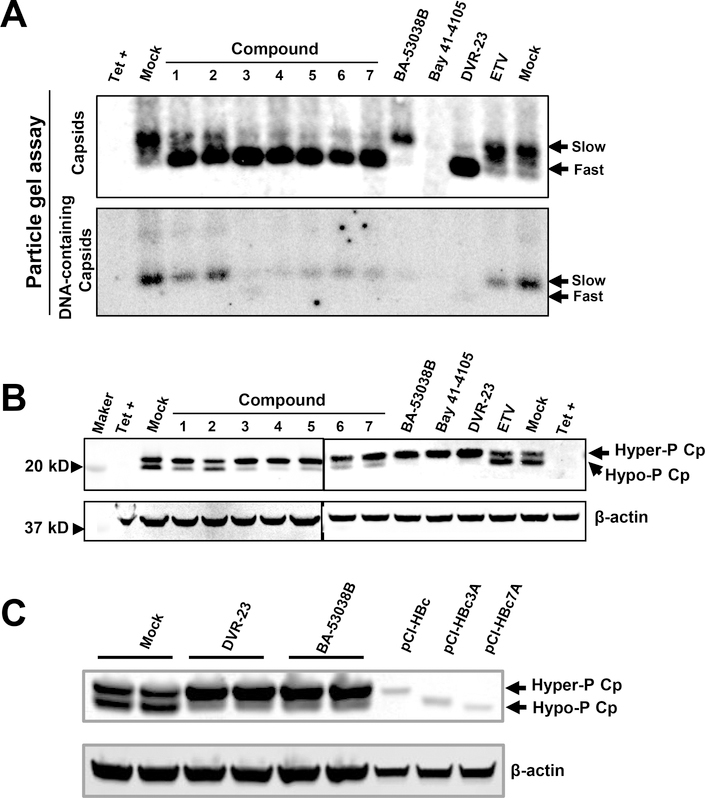
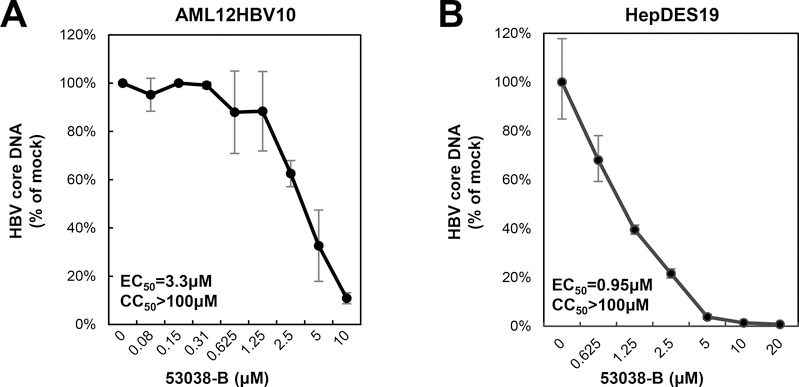
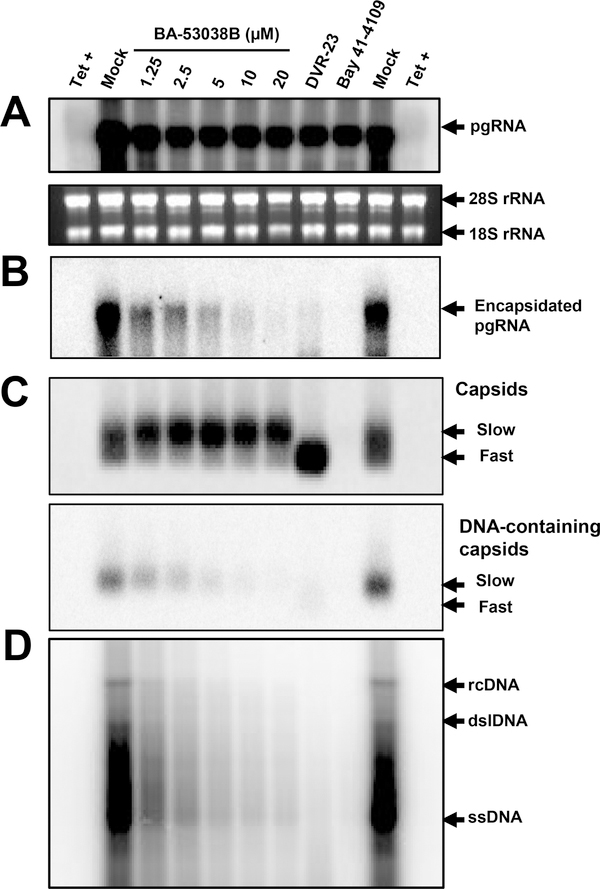
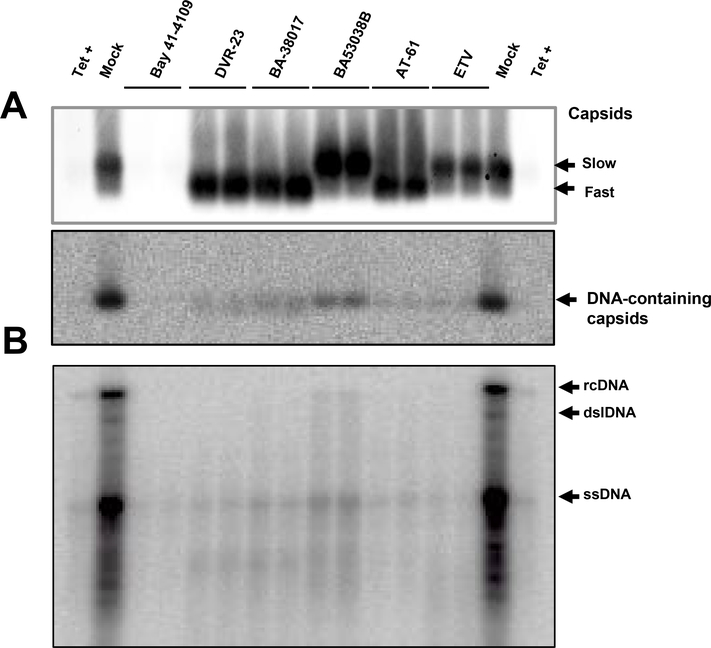
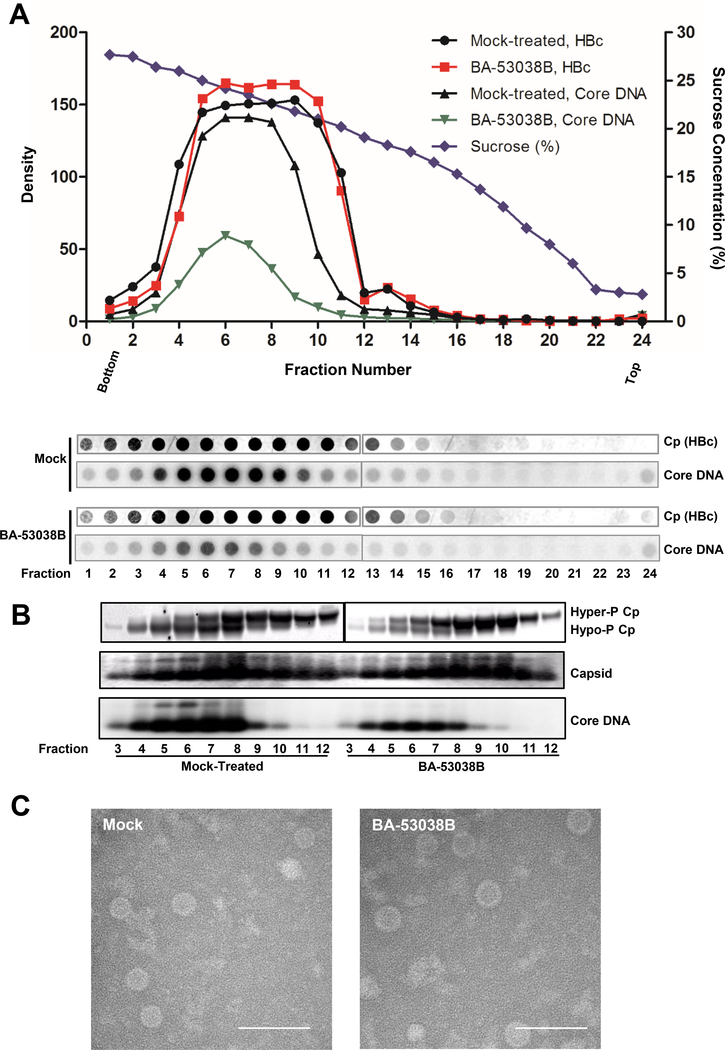
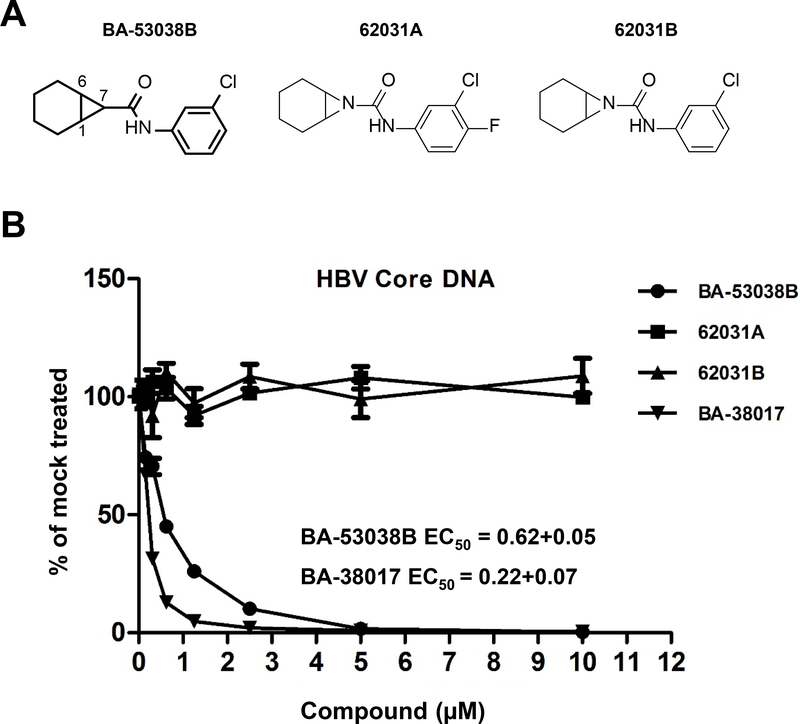
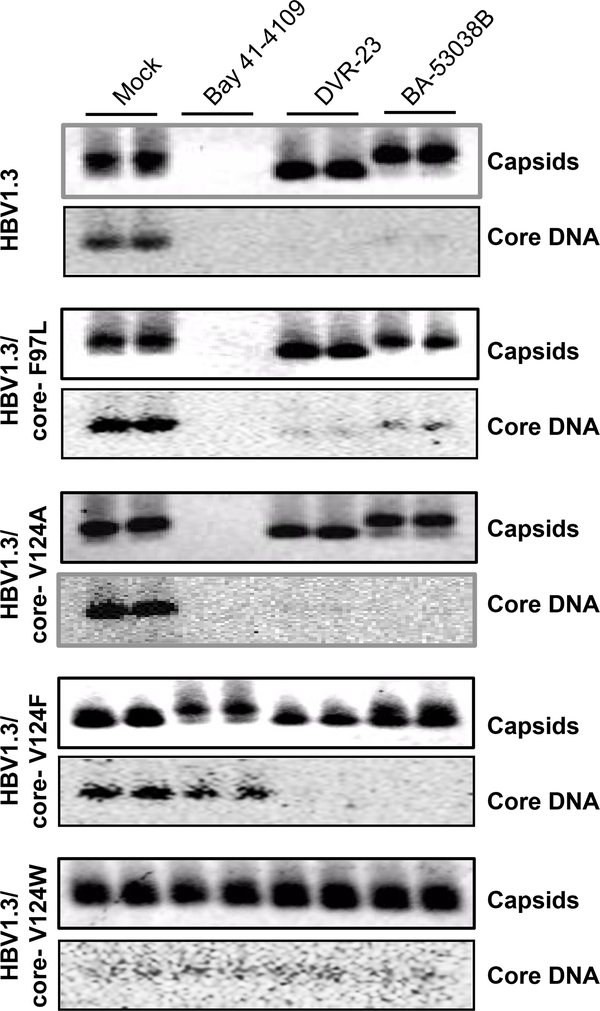
Hepatitis B virus (HBV) core protein is a small protein with 183 amino acid residues and assembles the pregenomic (pg) RNA and viral DNA polymerase to form nucleocapsids. During the last decades, several groups have reported HBV core protein allosteric modulators (CpAMs) with distinct chemical structures. CpAMs bind to the hydrophobic HAP pocket located at the dimer-dimer interface and induce allosteric conformational changes in the core protein subunits. While Type I CpAMs, heteroaryldihydropyrimidine (HAP) derivatives, misdirect core protein dimers to assemble noncapsid polymers, Type II CpAMs, represented by sulfamoylbenzamides, phenylpropenamides, and several other chemotypes, induce the assembly of empty capsids with global structural alterations and faster mobility in native agarose gel electrophoresis. Through high throughput screening of an Asinex small molecule library containing 19 920 compounds, we identified 8 structurally distinct CpAMs. While 7 of those compounds are typical Type II CpAMs, a novel benzamide derivative, designated as BA-53038B, induced the formation of morphologically "normal" empty capsids with slow electrophoresis mobility. Drug resistant profile analyses indicated that BA-53038B most likely bound to the HAP pocket but obviously modulated HBV capsid assembly in a distinct manner. BA-53038B and other CpAMs reported herein provide novel structure scaffolds for the development of core protein-targeted antiviral agents for the treatment of chronic hepatitis B.
Keywords: antiviral; capsid assembly; core protein allosteric modulator; hepatitis B virus; nucleocapsids.
Figures
References
-
- Mortalit GBD Causes of Death, C., Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death, 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 385 (9963), 117–71 DOI 10.1016/S0140-6736(14)61682-2. - DOI - PMC - PubMed
-
- Rehermann B; Ferrari C; Pasquinelli C; Chisari FV, The hepatitis B virus persists for decades after patients’ recovery from acute viral hepatitis despite active maintenance of a cytotoxic T- lymphocyte response. Nat Med 1996, 2 (10), 1104–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
