

Profiling proliferative cells and their progeny in damaged murine hearts
- PMID: 30530645
- PMCID: PMC6310797
- DOI: 10.1073/pnas.1805829115
Profiling proliferative cells and their progeny in damaged murine hearts
Abstract
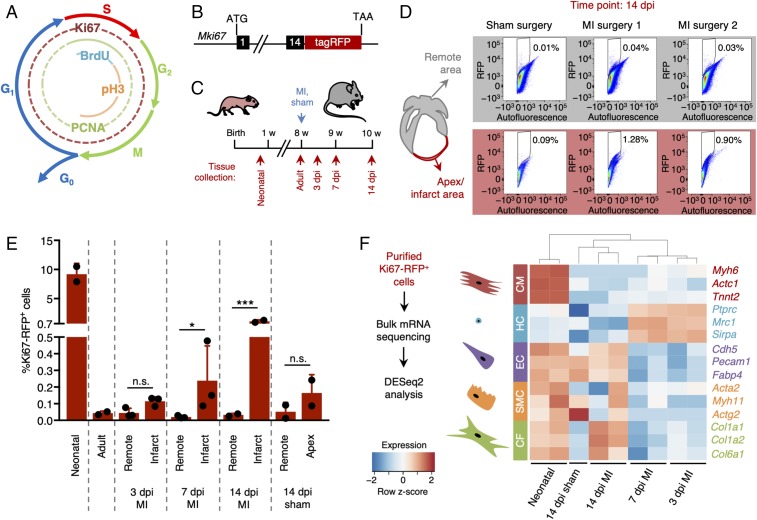
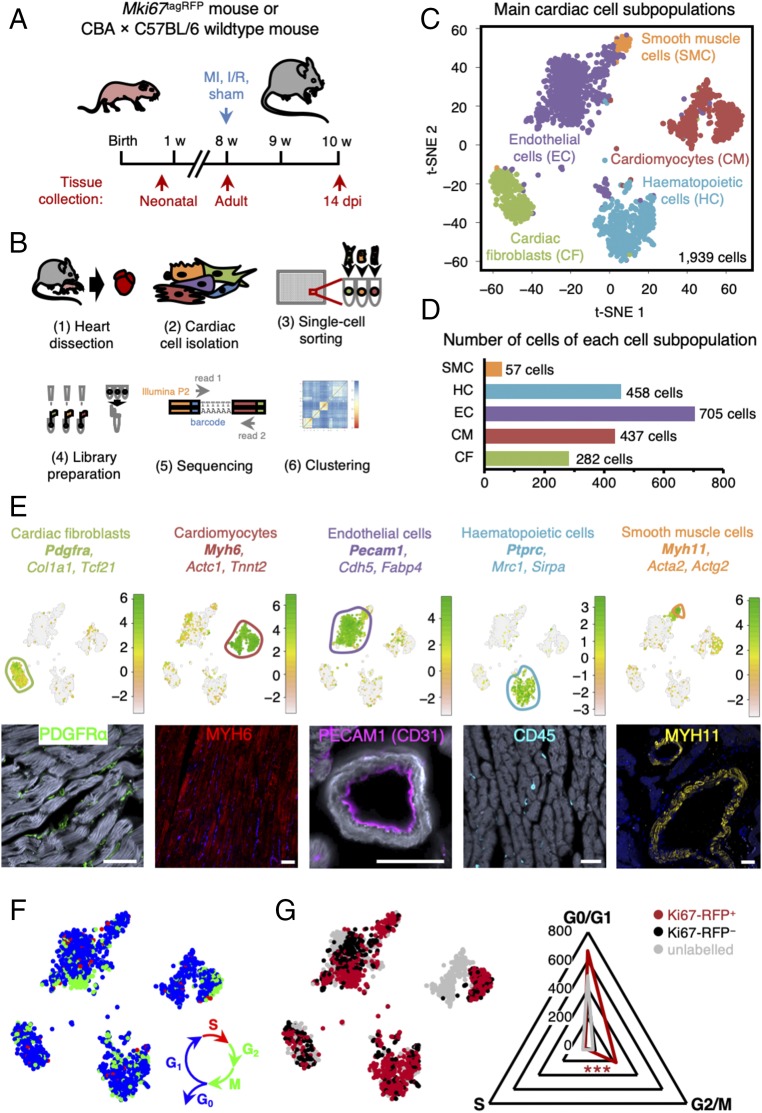
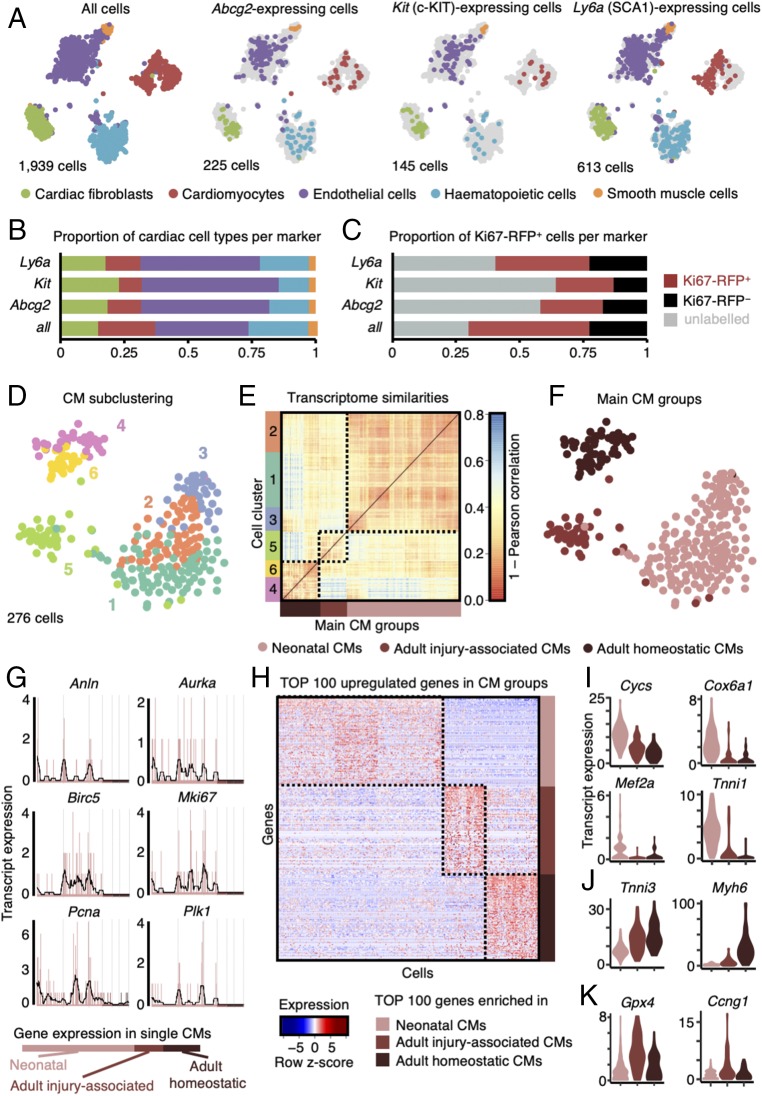
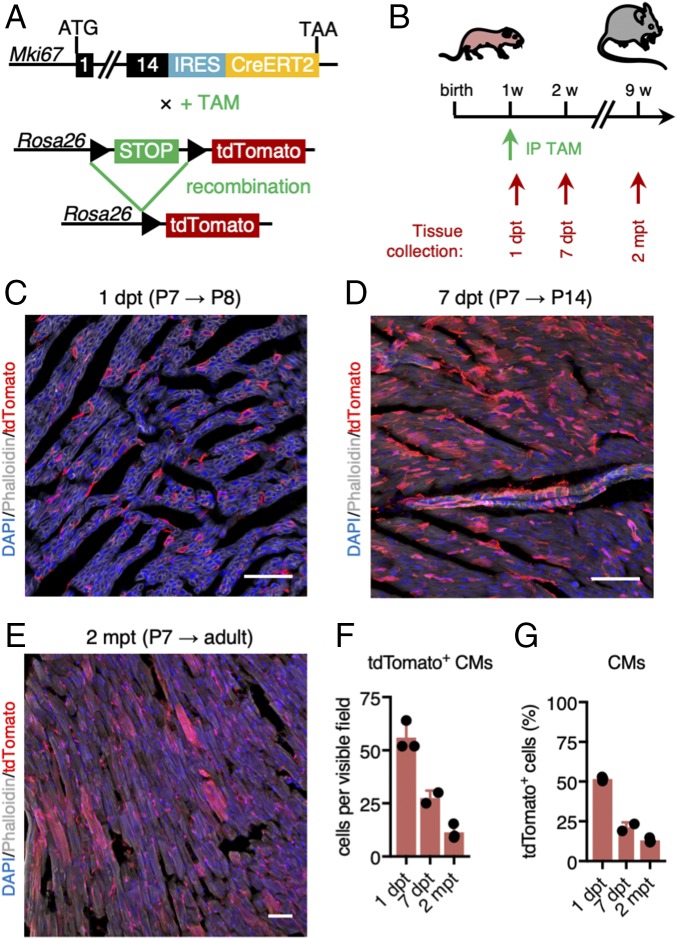
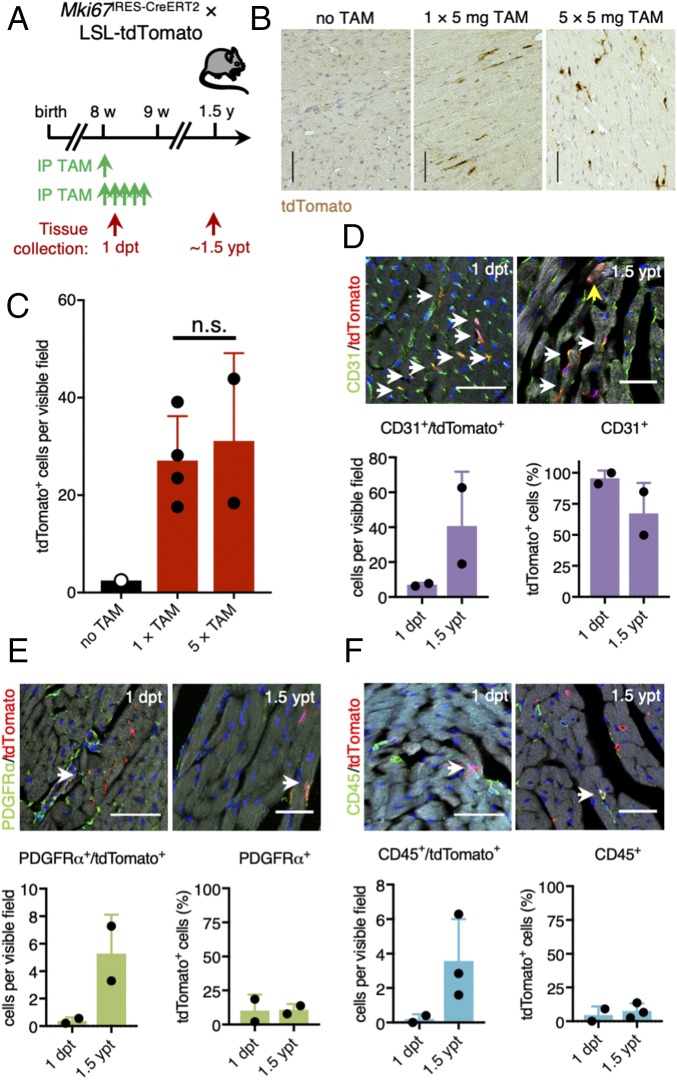
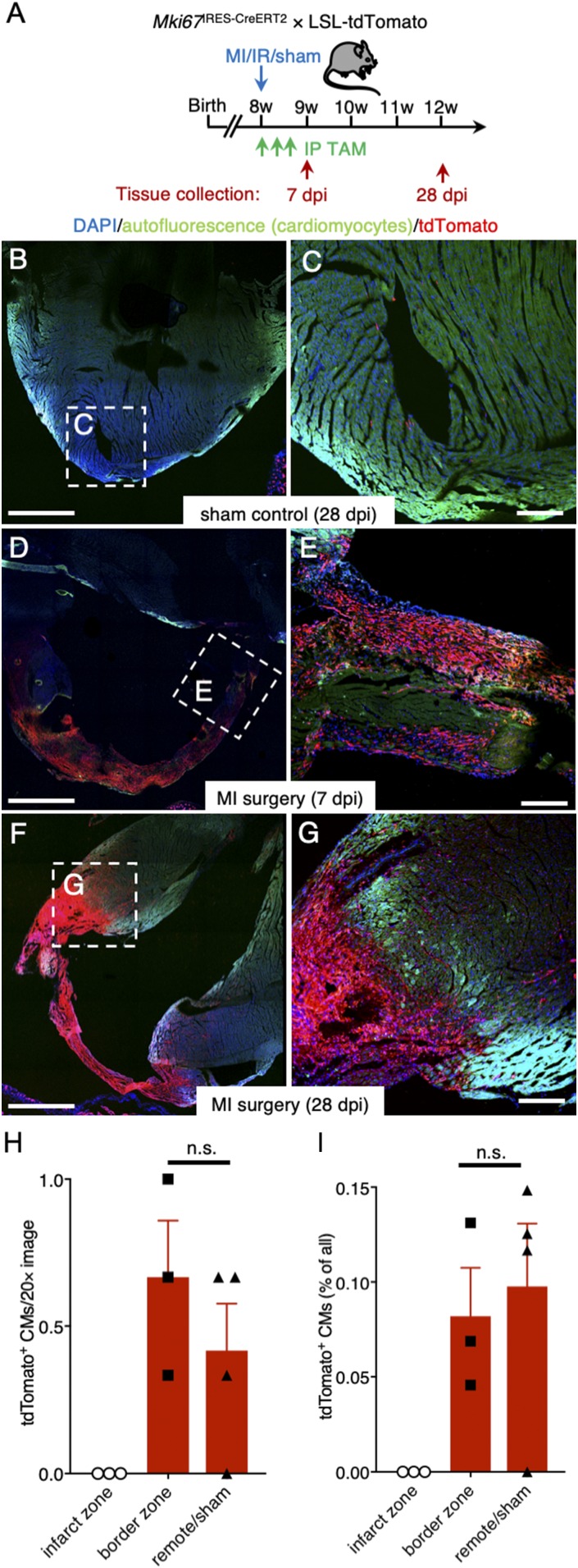
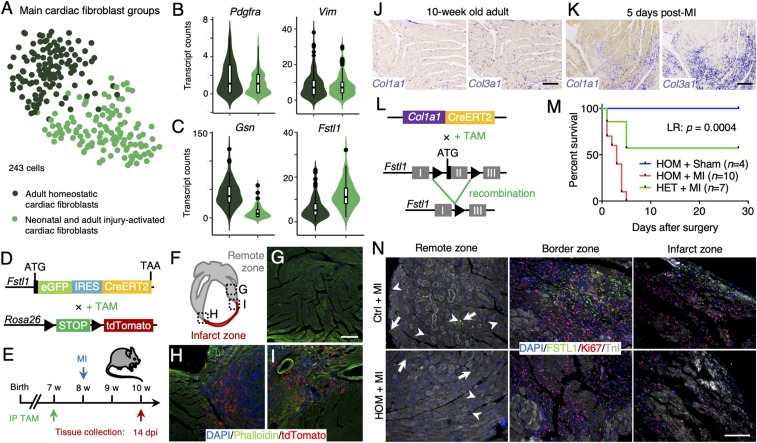
The significance of cardiac stem cell (CSC) populations for cardiac regeneration remains disputed. Here, we apply the most direct definition of stem cell function (the ability to replace lost tissue through cell division) to interrogate the existence of CSCs. By single-cell mRNA sequencing and genetic lineage tracing using two Ki67 knockin mouse models, we map all proliferating cells and their progeny in homoeostatic and regenerating murine hearts. Cycling cardiomyocytes were only robustly observed in the early postnatal growth phase, while cycling cells in homoeostatic and damaged adult myocardium represented various noncardiomyocyte cell types. Proliferative postdamage fibroblasts expressing follistatin-like protein 1 (FSTL1) closely resemble neonatal cardiac fibroblasts and form the fibrotic scar. Genetic deletion of Fstl1 in cardiac fibroblasts results in postdamage cardiac rupture. We find no evidence for the existence of a quiescent CSC population, for transdifferentiation of other cell types toward cardiomyocytes, or for proliferation of significant numbers of cardiomyocytes in response to cardiac injury.
Keywords: cardiac regeneration; fibroblasts; lineage tracing; single-cell transcriptomics; stem cells.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
Conflict of interest statement: H.C. is co-principal investigator (co-PI) on a Dutch grant on organs-on-a-chip with C.L.M. as co-PI, however, on an unrelated topic.
Figures
References
-
- Alkass K, et al. No evidence for cardiomyocyte number expansion in preadolescent mice. Cell. 2015;163:1026–1036. - PubMed
-
- Soonpaa MH, et al. Cardiomyocyte cell-cycle activity during preadolescence. Cell. 2015;163:781–782. - PubMed
-
- Beltrami AP, et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell. 2003;114:763–776. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
