

Estimating the number of sexual events per generation in a facultatively sexual haploid population
- PMID: 30531814
- PMCID: PMC6781114
- DOI: 10.1038/s41437-018-0171-1
Estimating the number of sexual events per generation in a facultatively sexual haploid population
Abstract
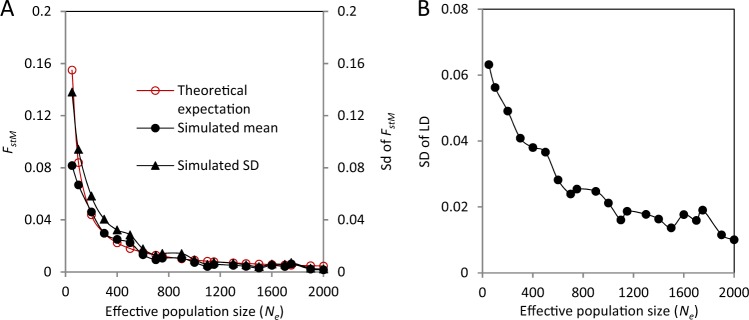
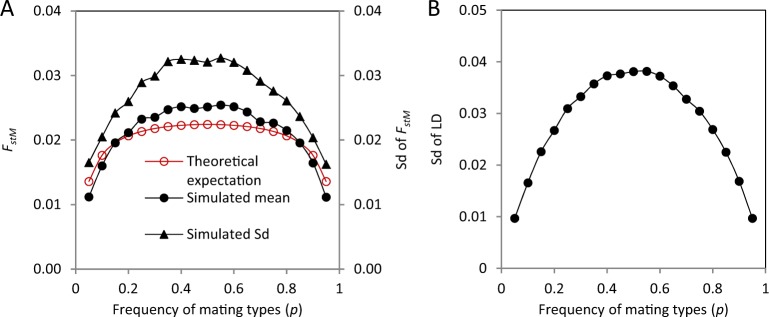
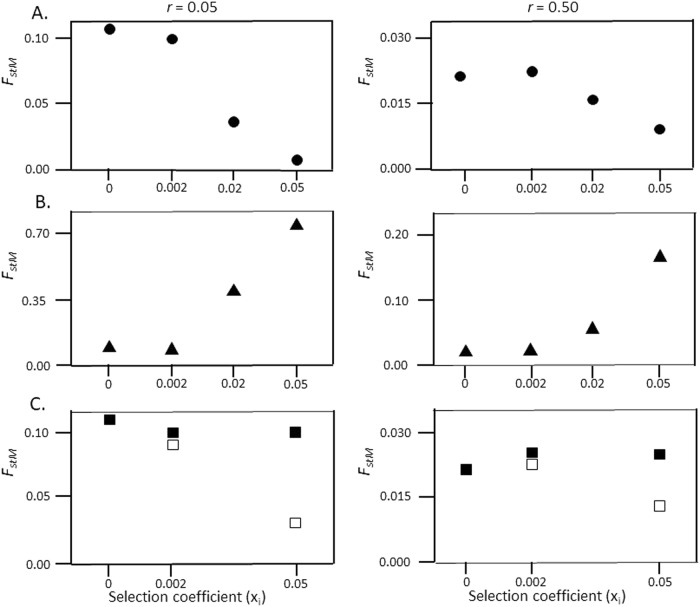
In populations of facultatively sexual organisms, the proportion of sexually produced offspring contributed to each generation is a critical determinant of their evolutionary potential. However, estimating this parameter in natural populations has proved difficult. Here we develop a population genetic model for estimating the number of sexual events occurring per generation for facultatively sexual haploids possessing a biallelic mating-type locus (e.g., Chlamydomonas, ascomycete fungi). Our model treats the population as two subpopulations possessing opposite mating-type alleles, which exchange genes only when a sexual event takes place. Where mating types are equally abundant, we show that, for a neutral genetic marker, genetic differentiation between mating-type subpopulations is a simple function of the effective population size, the frequency of sexual reproduction, and the recombination fraction between the genetic marker and the mating-type locus. We employ simulations to examine the effects of linkage of markers to the mating-type locus, inequality of mating-type frequencies, mutation rate, and selection on this relationship. Finally, we apply our model to estimate the number of sexual reproduction events per generation in populations of four species of facultatively sexual ascomycete fungi, which have been jointly scored for mating type and a range of polymorphic molecular markers. Relative estimates are in line with expectations based on the known reproductive biology of these species.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




References
-
- Agapow PM, Burt A. Indices of multilocus linkage disequilibrium. Mol Ecol Notes. 2001;1:101–102. doi: 10.1046/j.1471-8278.2000.00014.x. - DOI
-
- Ali S, Gladieux P, Rahman H, et al. Inferring the contribution of sexual reproduction, migration and off-season survival to the temporal maintenance of microbial populations: a case study on the wheat fungal pathogen Puccinia striiformis f.sp. tritici. Mol Ecol. 2014;23:603–617. doi: 10.1111/mec.12629. - DOI - PubMed
-
- Ali S, Soubeyrand S, Gladieux P, Giraud T, LeConte M, Gautier A, Mboup M, Chen W, De Vallavieille-Pope C, Enjalbert J, et al. CLONCASE: estimation of sex frequency and effective population size by clonemate resampling in partially clonal organisms. Mol Ecol Resour. 2016;16:845–861. doi: 10.1111/1755-0998.12511. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
