

Gas6 is dispensable for pubertal mammary gland development
- PMID: 30533018
- PMCID: PMC6289431
- DOI: 10.1371/journal.pone.0208550
Gas6 is dispensable for pubertal mammary gland development
Abstract
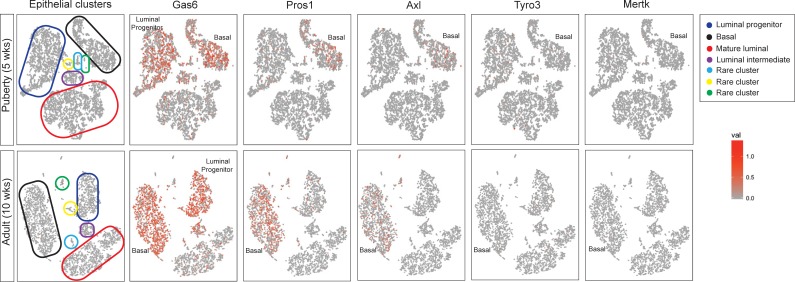
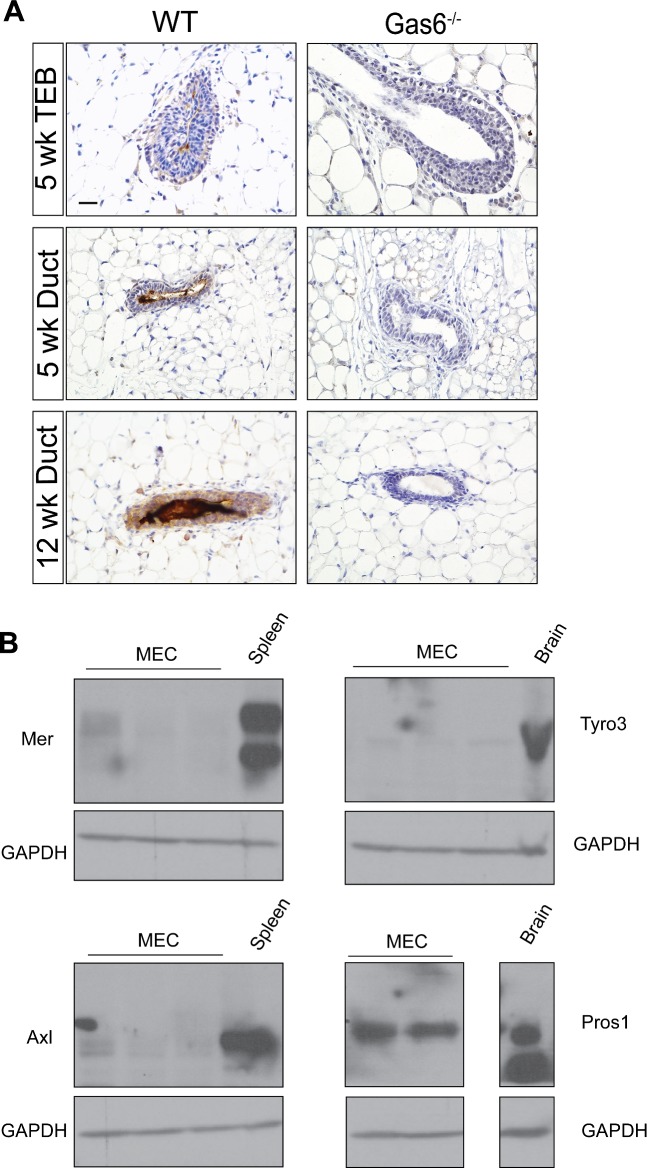
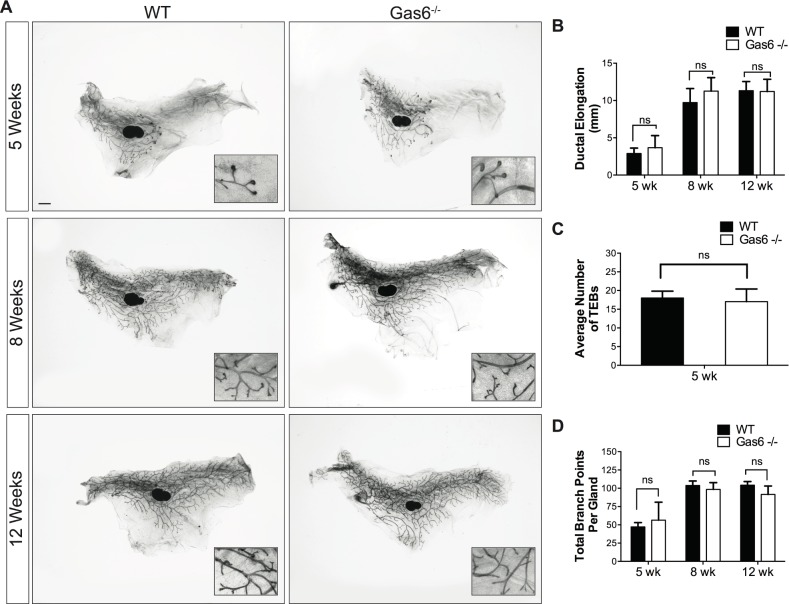
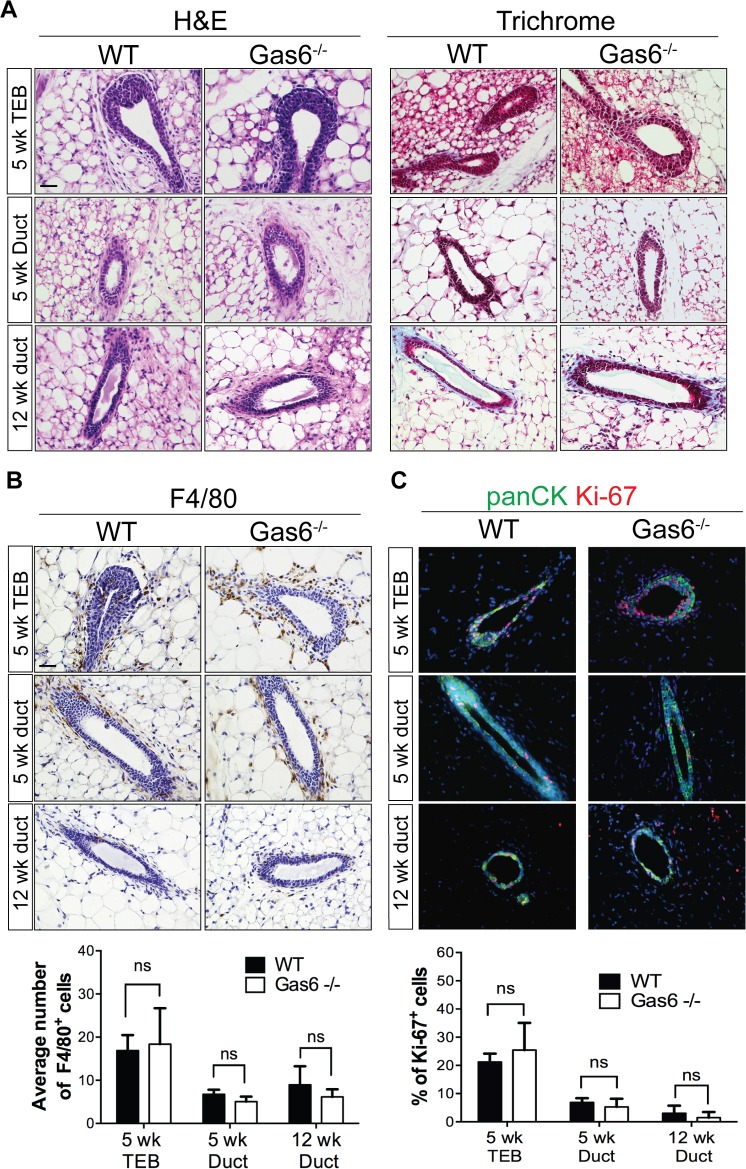
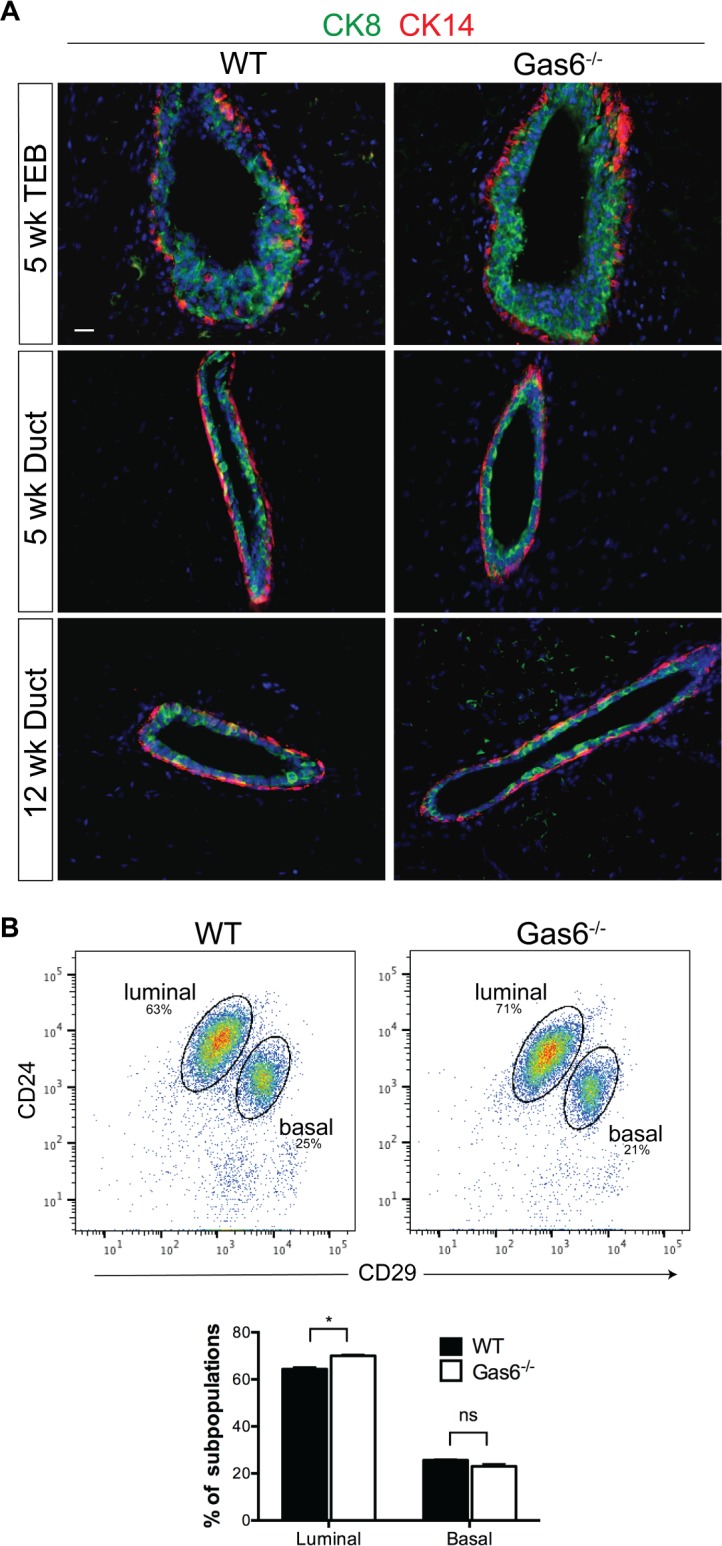
Mammary gland development is a complex and dynamic process that occurs mainly postnatally. Ductal elongation and branching morphogenesis are regulated by a plethora of factors, including cytokines, hormones, growth factors and the extracellular matrix. Gas6 is a secreted gamma-carboxylated protein that binds to a family of receptors tyrosine kinase receptors known as the TAMR family (Tyro3, Axl, Mer). Gas6 function in developmental processes has been shown in nervous, reproductive and immune systems. In this study, we found that Gas6 is highly expressed in virgin adult mammary glands but declines during pregnancy and lactation. Specifically, Gas6 is highly expressed in luminal and basal mammary epithelial cells during puberty and adulthood, while TAMR expression is low. Mammary whole mount analysis revealed that Gas6 germline deletion does not impact ductal elongation, branching morphogenesis or terminal end bud formation. Masson's trichrome staining showed that collagen deposition is similar in Gas6-/- mice as compared to wildtype mice. Gas6-/- mammary glands presented an organized luminal and myoepithelial bilayer of cells, and the proportion of mammary stem cells was unchanged in Gas6-/- mammary glands as compared to wildtype. Finally, proliferation of epithelial cells and macrophage number were similar in both groups. These studies suggest that Gas6 is not essential for pubertal mammary gland development in nulliparous mice.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Mailleux AA, Overholtzer M, Schmelzle T, Bouillet P, Strasser A, Brugge JS. BIM regulates apoptosis during mammary ductal morphogenesis, and its absence reveals alternative cell death mechanisms. Dev Cell. 2007;12(2):221–34. 10.1016/j.devcel.2006.12.003 ; PubMed Central PMCID: PMCPMC2698712. - DOI - PMC - PubMed
-
- Mailleux AA, Overholtzer M, Brugge JS. Lumen formation during mammary epithelial morphogenesis: insights from in vitro and in vivo models. Cell Cycle. 2008;7(1):57–62. Epub 2007/10/09. 10.4161/cc.7.1.5150 . - DOI - PubMed
-
- Humphreys RC, Krajewska M, Krnacik S, Jaeger R, Weiher H, Krajewski S, et al. Apoptosis in the terminal endbud of the murine mammary gland: a mechanism of ductal morphogenesis. Development. 1996;122(12):4013–22. . - PubMed
-
- Stitt TN, Conn G, Gore M, Lai C, Bruno J, Radziejewski C, et al. The anticoagulation factor protein S and its relative, Gas6, are ligands for the Tyro 3/Axl family of receptor tyrosine kinases. Cell. 1995;80(4):661–70. . - PubMed
-
- Lemke G. Biology of the TAM receptors. Cold Spring Harb Perspect Biol. 2013;5(11):a009076 10.1101/cshperspect.a009076 ; PubMed Central PMCID: PMCPMC3809585. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
