

Non-coding RNA in C9orf72-related amyotrophic lateral sclerosis and frontotemporal dementia: A perfect storm of dysfunction
- PMID: 30533567
- PMCID: PMC6260478
- DOI: 10.1016/j.ncrna.2018.09.001
Non-coding RNA in C9orf72-related amyotrophic lateral sclerosis and frontotemporal dementia: A perfect storm of dysfunction
Erratum in
-
Erratum regarding previously published articles.Noncoding RNA Res. 2020 Nov 7;5(4):219. doi: 10.1016/j.ncrna.2020.11.001. eCollection 2020 Dec. Noncoding RNA Res. 2020. PMID: 33294747 Free PMC article.
Abstract
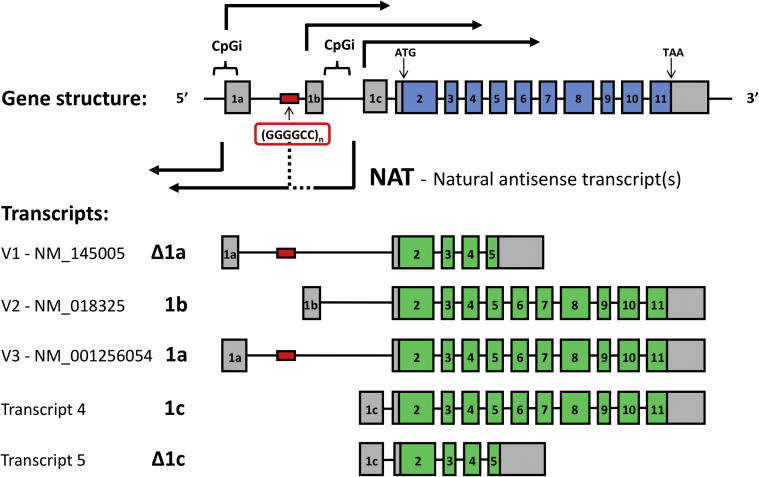
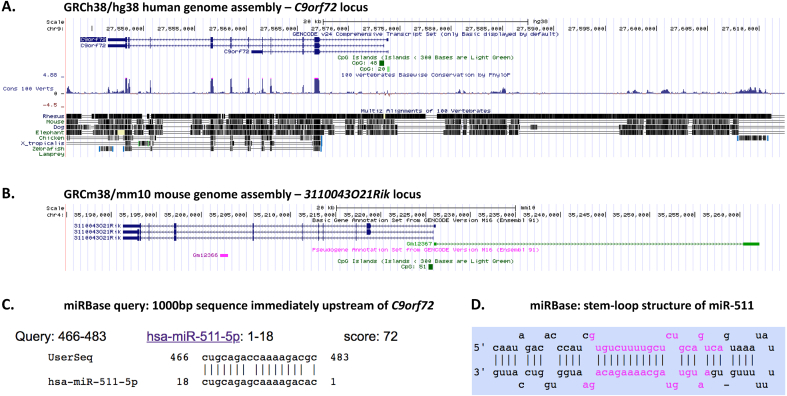
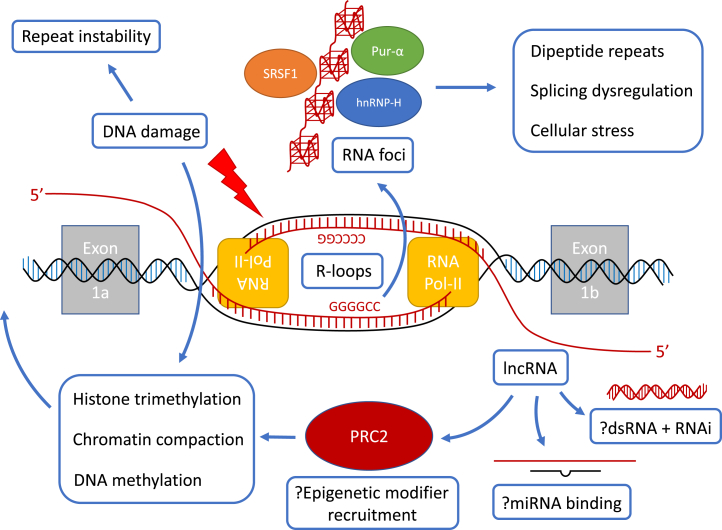
A hexanucleotide repeat expansion in the first intron/promoter region of C9orf72 is the most common genetic cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Both sense and antisense transcripts exist at the C9orf72 locus but the function of the antisense lncRNA is unknown. RNA toxicity of the transcribed repeat expansion has been implicated in the pathogenesis of C9orf72-related ALS/FTD, not only through direct sequestration of important RNA binding proteins but also indirectly through non-ATG dependent translation into dipeptide repeats. Formation of RNA/DNA hybrid R-loops may also play a key role in the pathogenesis of this condition and this mechanism could provide a link between the repeat expansion, DNA damage, repeat instability and deficiency of RNA binding proteins. Non-coding C9orf72 antisense transcripts could also act to epigenetically regulate gene expression at the locus. The potential effects of such non-coding RNAs should be considered in the design of antisense oligonucleotide therapeutics for C9orf72-related ALS/FTD. Furthermore, the mechanisms of RNA dysregulation exemplified by C9orf72-related disease may help illustrate more broadly how a "perfect storm" of dysfunction occurs in ALS/FTD and how targeting these factors could lead to corrective or preventative therapies.
Keywords: ALS; Amyotrophic lateral sclerosis; C9orf72; FTD; Frontotemporal dementia; Non-coding RNA.
Figures
References
-
- Brown R.H., Al-Chalabi A. Amyotrophic lateral sclerosis. N. Engl. J. Med. 2017;377:162–172. - PubMed
-
- Byrne S., Walsh C., Lynch C., Bede P., Elamin M., Kenna K., Mclaughlin R., Hardiman O. Rate of familial amyotrophic lateral sclerosis: a systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry. 2011;82:623–627. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
