

Cellular RNA Helicase DHX9 Interacts with the Essential Epstein-Barr Virus (EBV) Protein SM and Restricts EBV Lytic Replication
- PMID: 30541834
- PMCID: PMC6364004
- DOI: 10.1128/JVI.01244-18
Cellular RNA Helicase DHX9 Interacts with the Essential Epstein-Barr Virus (EBV) Protein SM and Restricts EBV Lytic Replication
Abstract
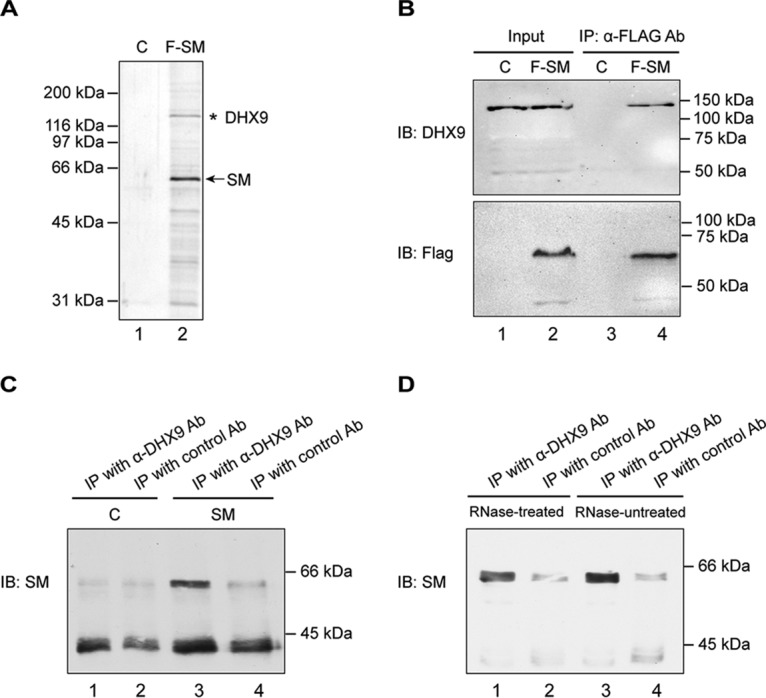
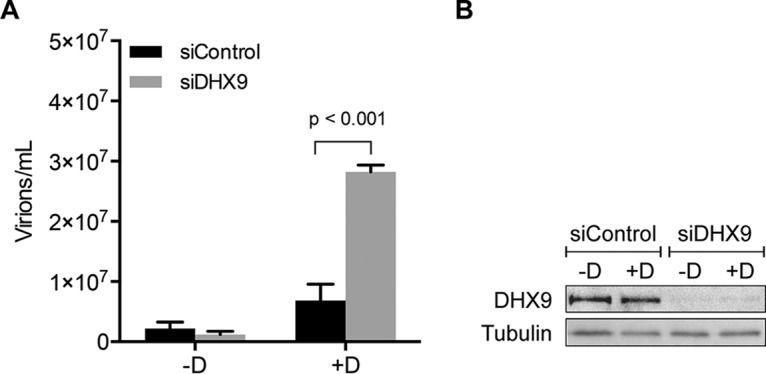
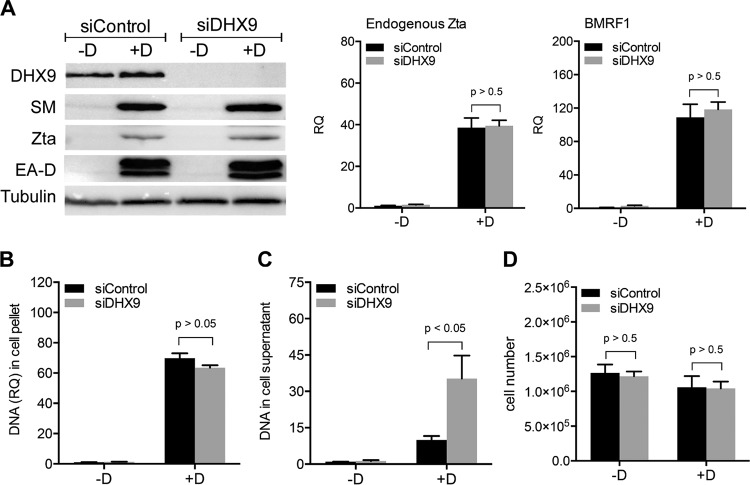
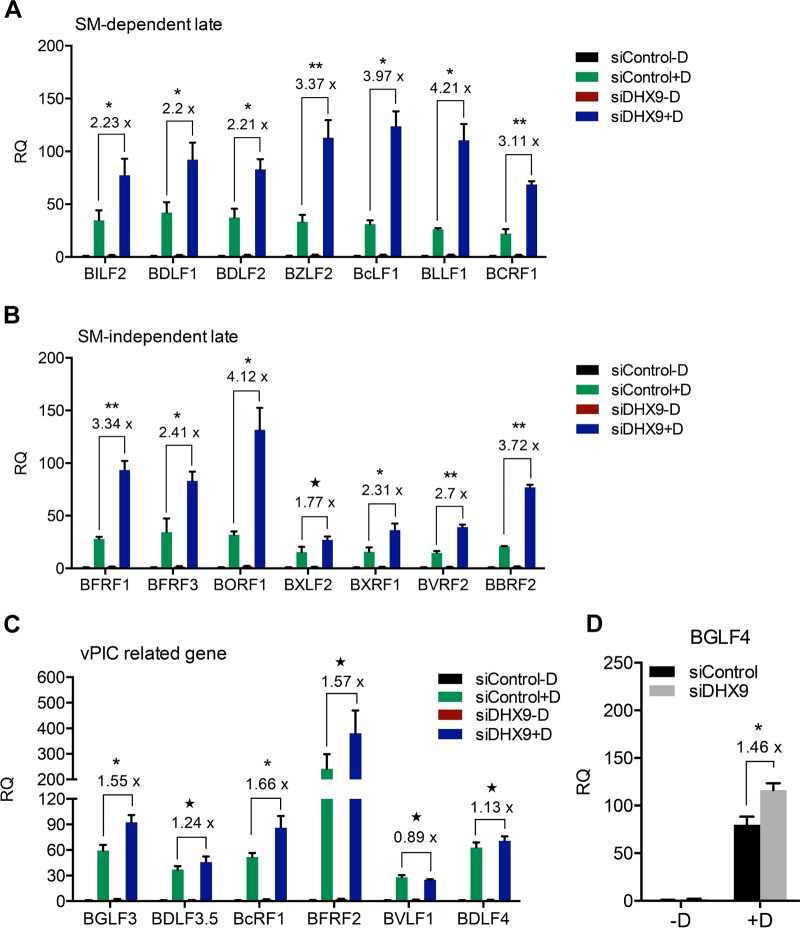
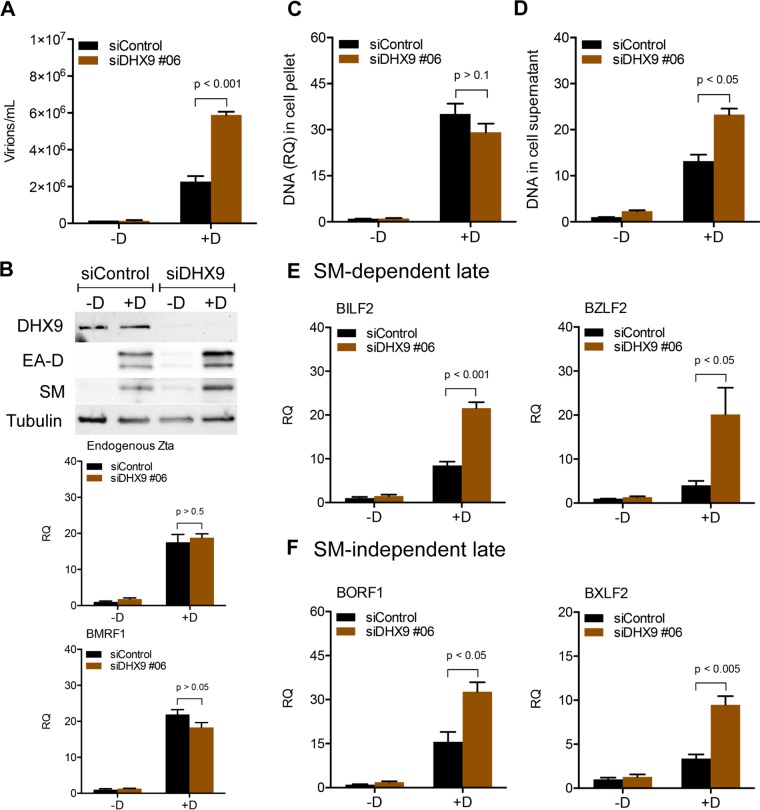
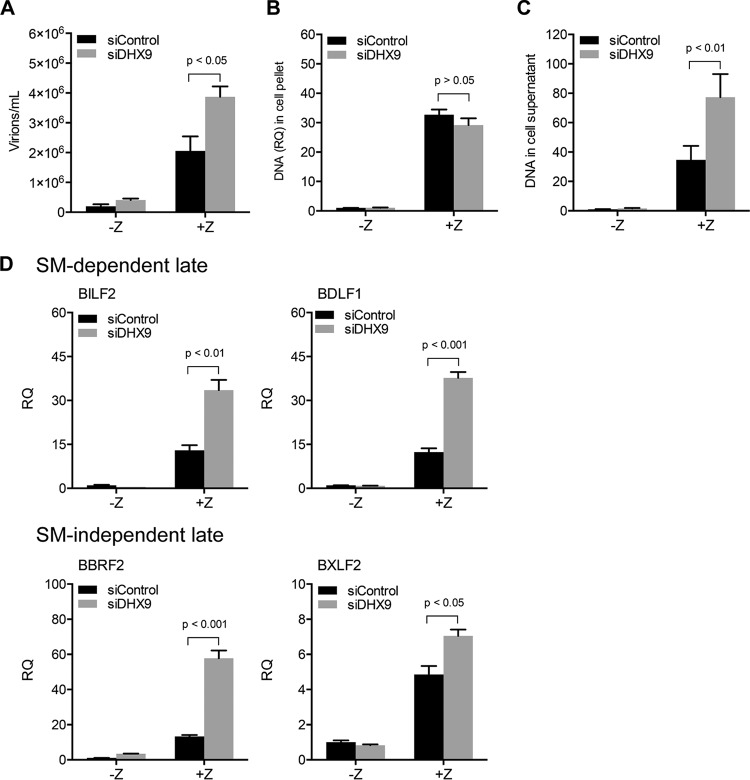
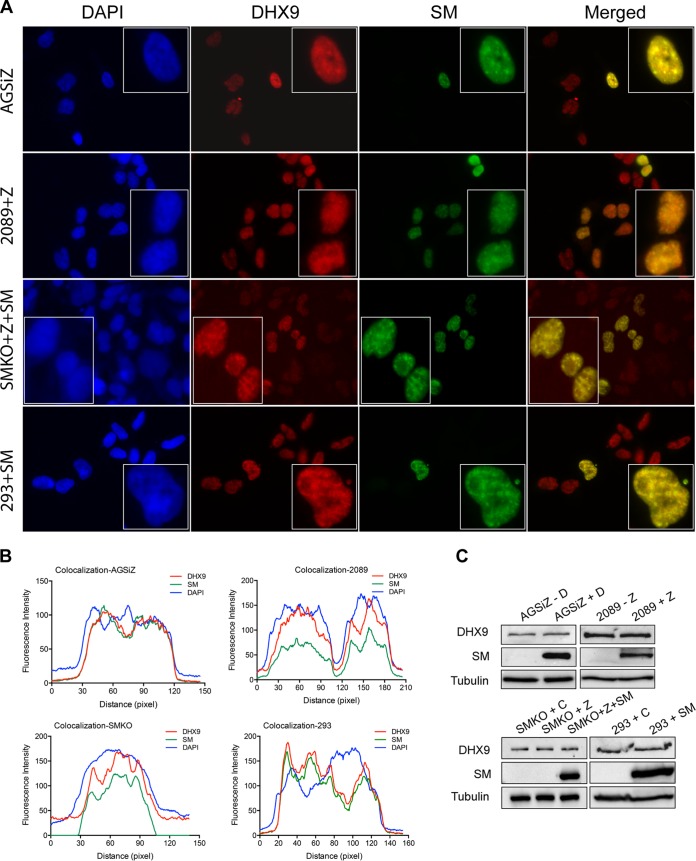
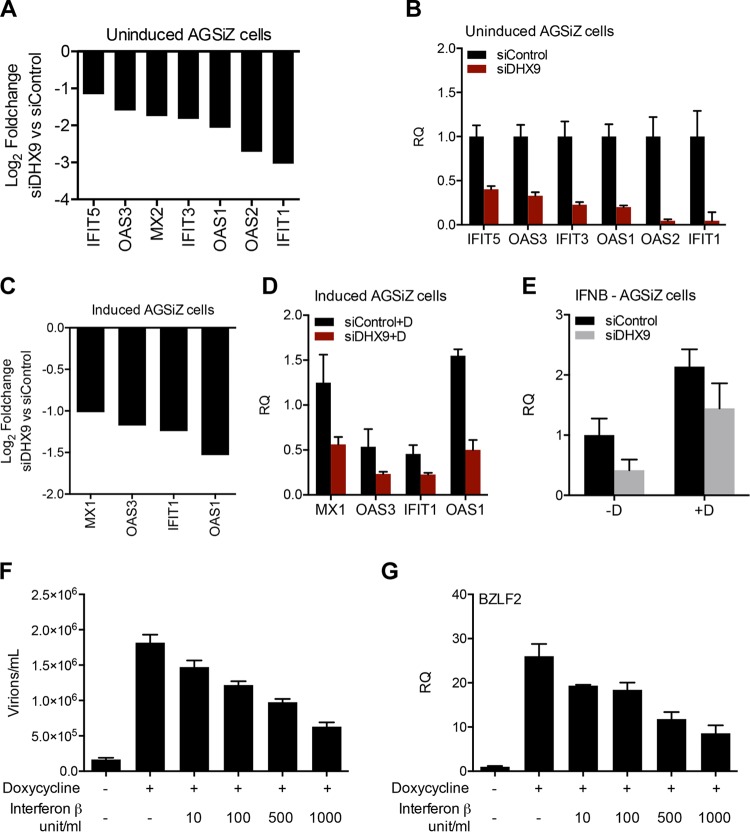
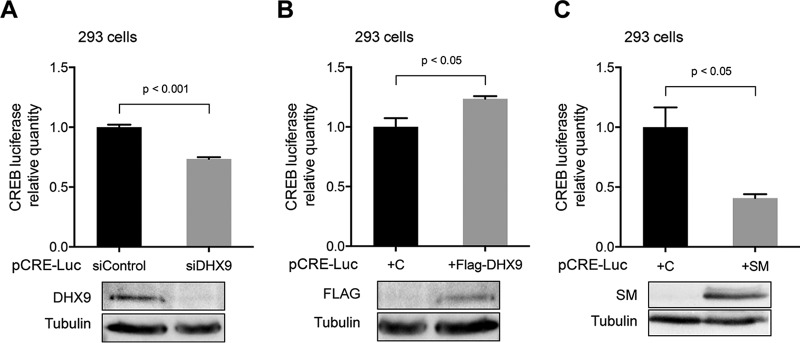
Epstein-Barr virus (EBV) SM protein is an RNA-binding protein that has multiple posttranscriptional gene regulatory functions essential for EBV lytic replication. In this study, we identified an interaction between SM and DHX9, a DExH-box helicase family member, by mass spectrometry and coimmunoprecipitation. DHX9 participates in many cellular pathways involving RNA, including transcription, processing, transport, and translation. DHX9 enhances virus production or infectivity of a wide variety of DNA and RNA viruses. Surprisingly, an increase in EBV late gene expression and virion production occurred upon knockdown of DHX9. To further characterize the SM-DHX9 interaction, we performed immunofluorescence microscopy of EBV-infected cells and found that DHX9 partially colocalized with SM in nuclear foci during EBV lytic replication. However, the positive effect of DHX9 depletion on EBV lytic gene expression was not confined to SM-dependent genes, indicating that the antiviral effect of DHX9 was not mediated through its effects on SM. DHX9 enhanced activation of innate antiviral pathways comprised of several interferon-stimulated genes that are active against EBV. SM inhibited the transcription-activating function of DHX9, which acts through cAMP response elements (CREs), suggesting that SM may also act to counteract DHX9's antiviral functions during lytic replication.IMPORTANCE This study identifies an interaction between Epstein-Barr virus (EBV) SM protein and cellular helicase DHX9, exploring the roles that this interaction plays in viral infection and host defenses. Whereas most previous studies established DHX9 as a proviral factor, we demonstrate that DHX9 may act as an inhibitor of EBV virion production. DHX9 enhanced innate antiviral pathways active against EBV and was needed for maximal expression of several interferon-induced genes. We show that SM binds to and colocalizes DHX9 and may counteract the antiviral function of DHX9. These data indicate that DHX9 possesses antiviral activity and that SM may suppress the antiviral functions of DHX9 through this association. Our study presents a novel host-pathogen interaction between EBV and the host cell.
Keywords: Epstein-Barr virus; RNA processing; helicase; innate immune response; protein-protein interactions; transcriptional activation; virus-host interactions.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
The SWI/SNF Chromatin Regulator BRG1 Modulates the Transcriptional Regulatory Activity of the Epstein-Barr Virus DNA Polymerase Processivity Factor BMRF1.J Virol. 2017 Apr 13;91(9):e02114-16. doi: 10.1128/JVI.02114-16. Print 2017 May 1. J Virol. 2017. PMID: 28228591 Free PMC article.
-
Epstein-Barr virus co-opts TFIIH component XPB to specifically activate essential viral lytic promoters.Proc Natl Acad Sci U S A. 2020 Jun 9;117(23):13044-13055. doi: 10.1073/pnas.2000625117. Epub 2020 May 20. Proc Natl Acad Sci U S A. 2020. PMID: 32434920 Free PMC article.
-
Efficient Translation of Epstein-Barr Virus (EBV) DNA Polymerase Contributes to the Enhanced Lytic Replication Phenotype of M81 EBV.J Virol. 2018 Feb 26;92(6):e01794-17. doi: 10.1128/JVI.01794-17. Print 2018 Mar 15. J Virol. 2018. PMID: 29263273 Free PMC article.
-
RNA helicase A as co-factor for DNA viruses during replication.Virus Res. 2021 Jan 2;291:198206. doi: 10.1016/j.virusres.2020.198206. Epub 2020 Oct 23. Virus Res. 2021. PMID: 33132162 Review.
-
Latent and lytic Epstein-Barr virus replication strategies.Rev Med Virol. 2005 Jan-Feb;15(1):3-15. doi: 10.1002/rmv.441. Rev Med Virol. 2005. PMID: 15386591 Review.
Cited by
-
Awakening the sleeping giant: Epstein-Barr virus reactivation by biological agents.Pathog Dis. 2024 Feb 7;82:ftae002. doi: 10.1093/femspd/ftae002. Pathog Dis. 2024. PMID: 38281067 Free PMC article. Review.
-
The pseudogene GBP1P1 suppresses influenza A virus replication by acting as a protein decoy for DHX9.J Virol. 2024 Jul 23;98(7):e0073824. doi: 10.1128/jvi.00738-24. Epub 2024 Jun 28. J Virol. 2024. PMID: 38940585 Free PMC article.
-
Nasopharyngeal Carcinoma: The Role of the EGFR in Epstein-Barr Virus Infection.Pathogens. 2021 Aug 31;10(9):1113. doi: 10.3390/pathogens10091113. Pathogens. 2021. PMID: 34578147 Free PMC article. Review.
-
Identifying the key regulators orchestrating Epstein-Barr virus reactivation.Front Microbiol. 2024 Dec 5;15:1505191. doi: 10.3389/fmicb.2024.1505191. eCollection 2024. Front Microbiol. 2024. PMID: 39703703 Free PMC article. Review.
-
The SARS-CoV-2 helicase as a target for antiviral therapy: Identification of potential small molecule inhibitors by in silico modelling.J Mol Graph Model. 2022 Jul;114:108193. doi: 10.1016/j.jmgm.2022.108193. Epub 2022 Apr 18. J Mol Graph Model. 2022. PMID: 35462185 Free PMC article.
References
-
- Longnecker RM, Kieff E, Cohen JI. 2013. Epstein-Barr virus, p 1898–1959. In Knipe DM, Howley PM, Cohen JL, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed, vol 2 Lippincott Williams & Wilkins, Philadelphia, PA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
