

The Triticum Mosaic Virus Internal Ribosome Entry Site Relies on a Picornavirus-Like YX-AUG Motif To Designate the Preferred Translation Initiation Site and To Likely Target the 18S rRNA
- PMID: 30541835
- PMCID: PMC6384068
- DOI: 10.1128/JVI.01705-18
The Triticum Mosaic Virus Internal Ribosome Entry Site Relies on a Picornavirus-Like YX-AUG Motif To Designate the Preferred Translation Initiation Site and To Likely Target the 18S rRNA
Abstract
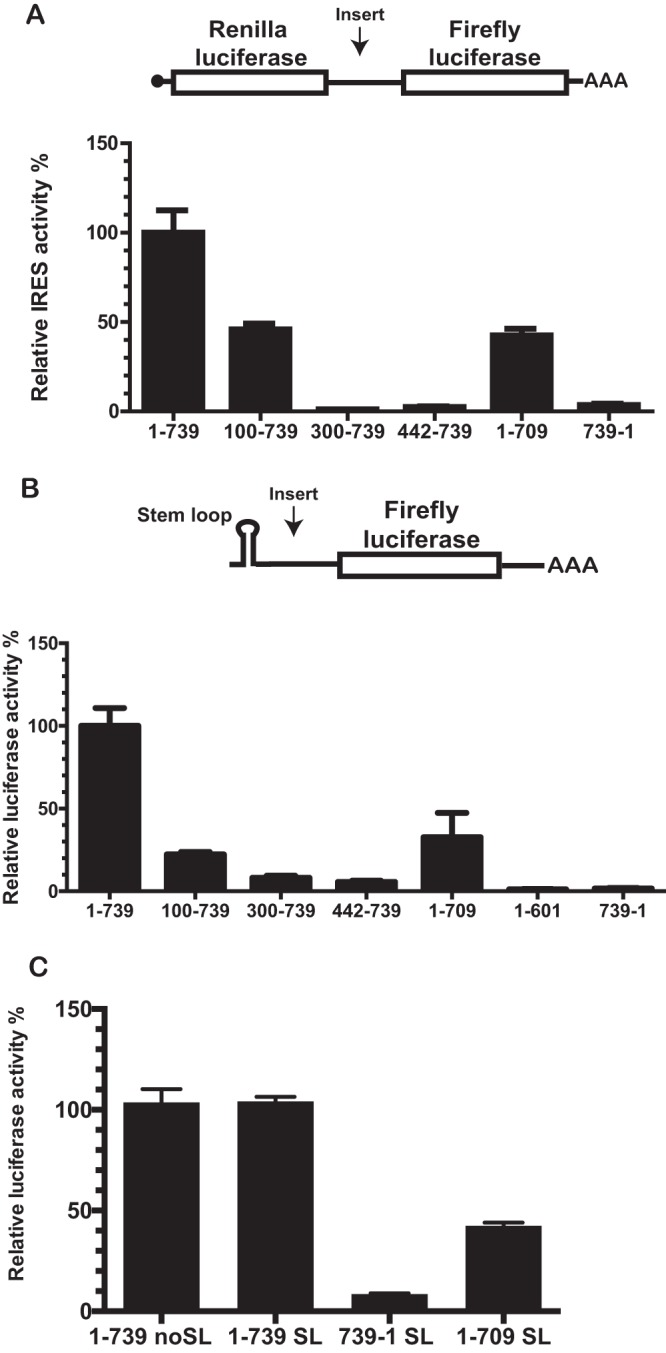
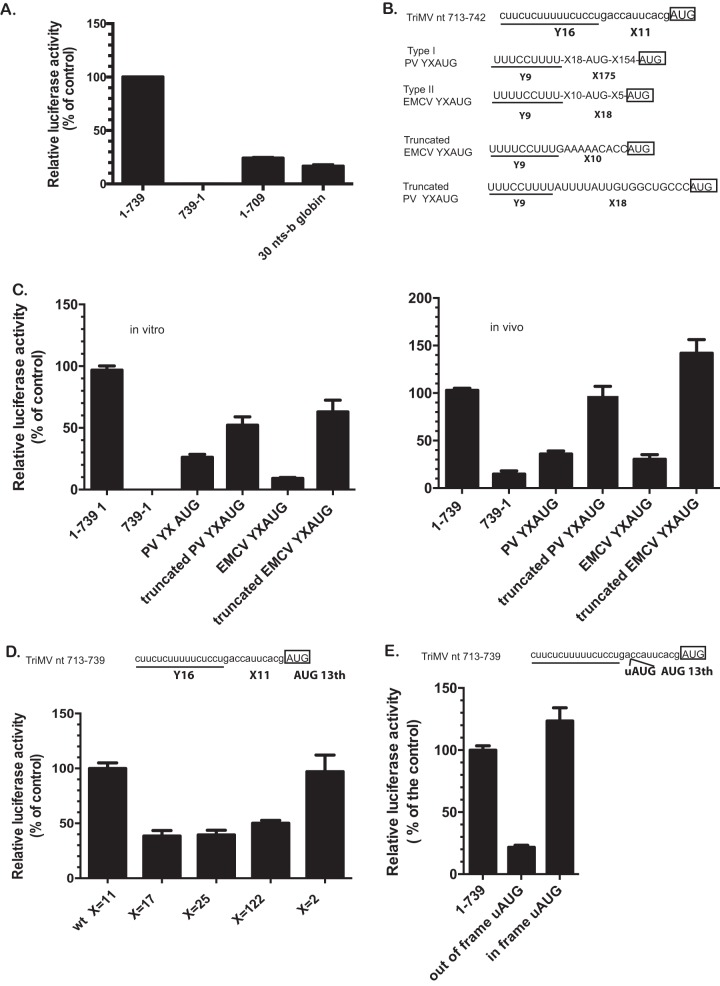
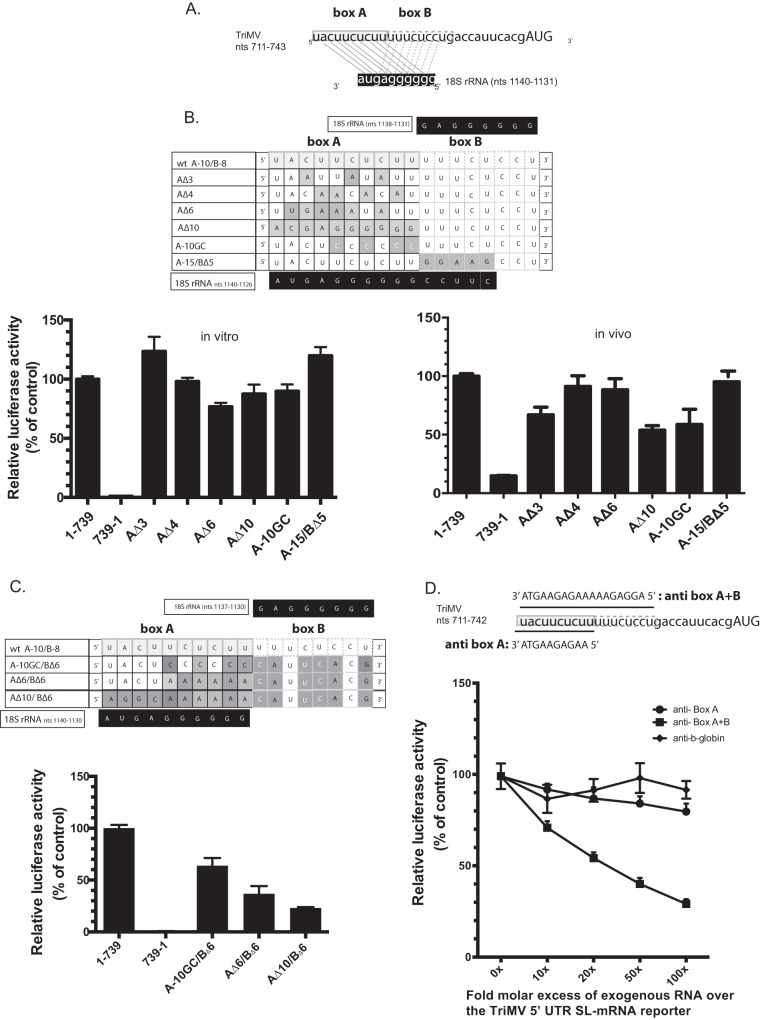
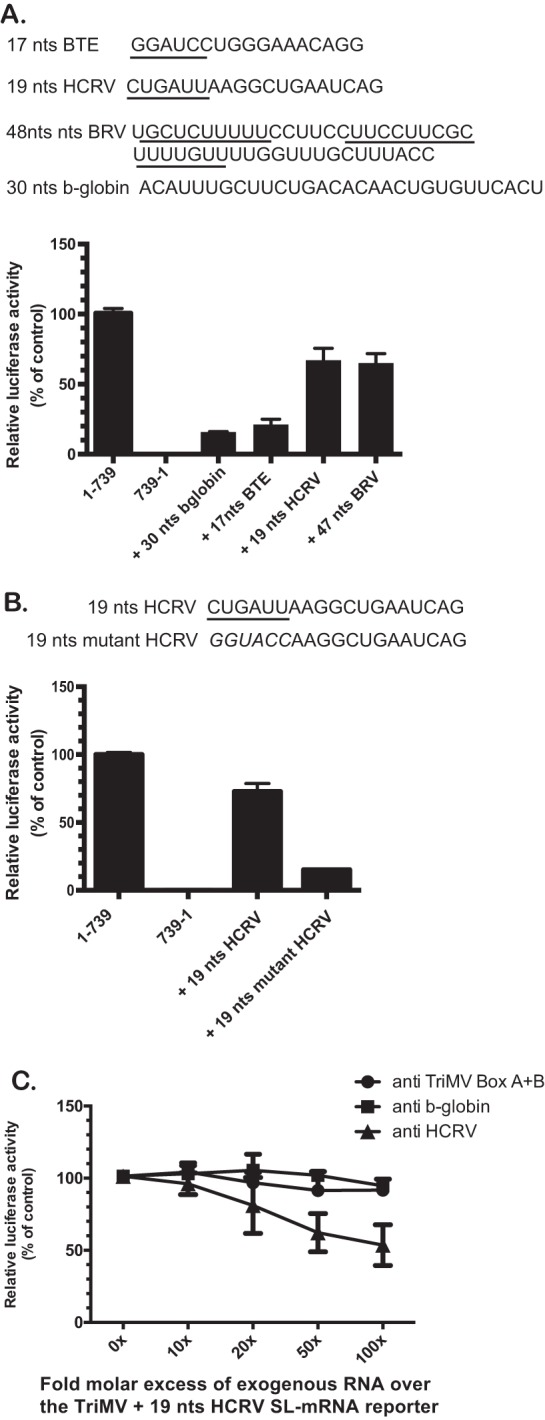
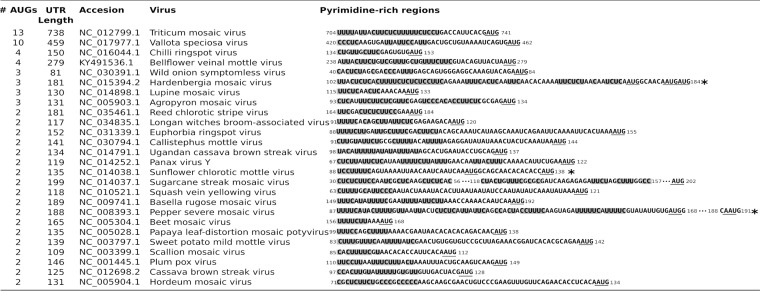
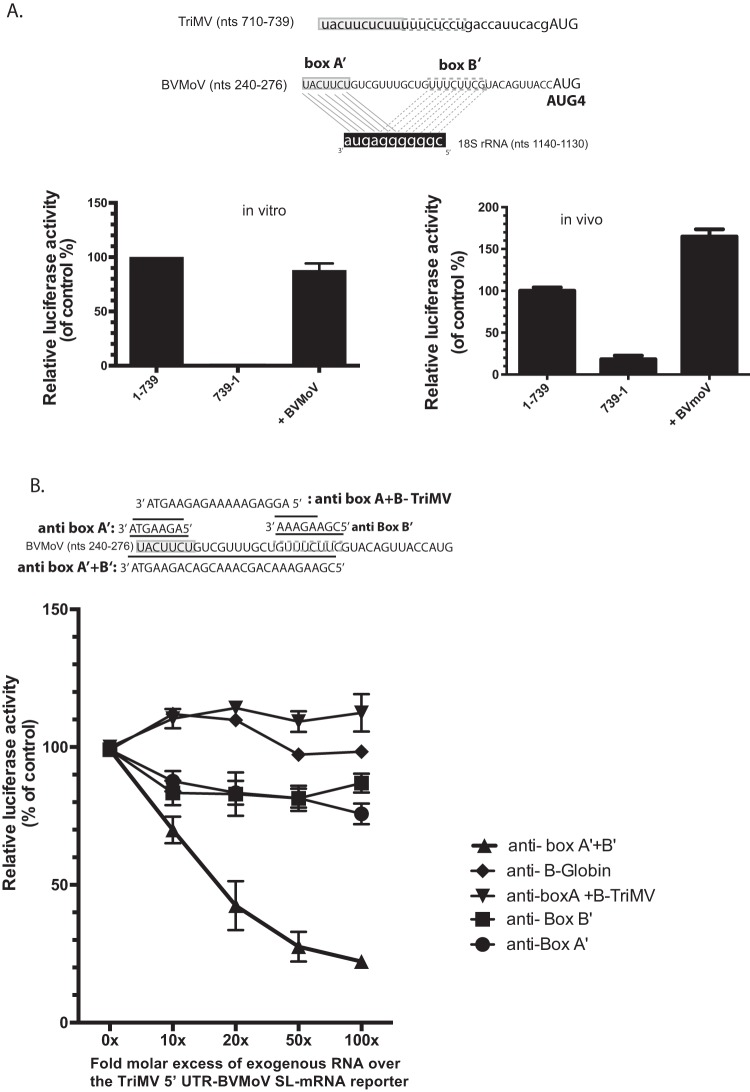
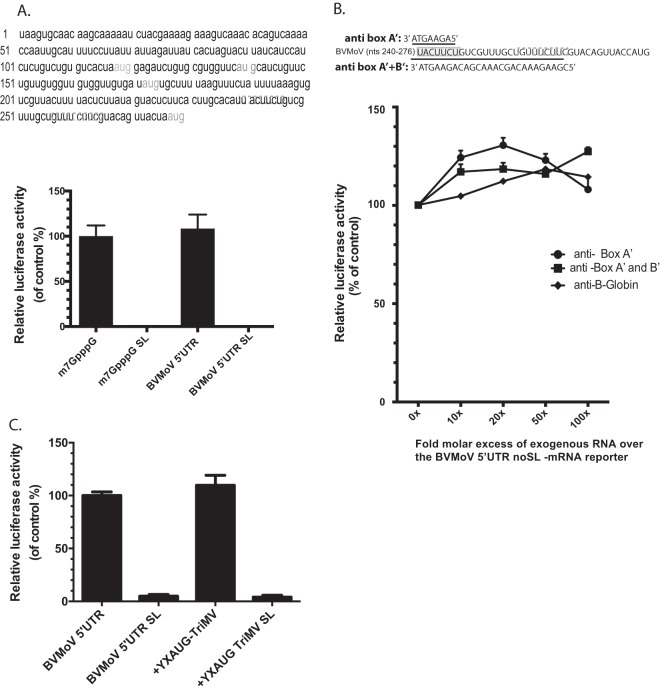
Several viruses encode an internal ribosome entry site (IRES) at the 5' end of their RNA, which, unlike most cellular mRNAs, initiates translation in the absence of a 5' m7GpppG cap. Here, we report a uniquely regulated translation enhancer found in the 739-nucelotide (nt) sequence of the Triticum mosaic virus (TriMV) leader sequence that distinguishes the preferred initiation site from a plethora of IRES-encoded AUG triplets. Through deletion mutations of the TriMV 5' untranslated region (UTR), we show that the TriMV 5' UTR encodes a cis-acting picornaviral Y16-X11-AUG-like motif with a 16-nt polypyrimidine CU-tract (Y16), at a precise, 11-nt distance (X11) from the preferred 13th AUG. Phylogenetic analyses indicate that this motif is conserved among potyviral leader sequences with multiple AUGs. Consistent with a broadly conserved mechanism, the motif could be functionally replaced with known picornavirus YX-AUG motifs and is predicted to function as target sites for the 18S rRNA by direct base pairing. Accordingly, mutations that disrupted overall complementarity to the 18S rRNA markedly reduced TriMV IRES activity, as did the delivery of antisense oligonucleotides designed to block YX-AUG accessibility. To our knowledge, this is the first report of a plant viral IRES YX-AUG motif, and our findings suggest that a conserved mechanism regulates translation for multiple economically important plant and animal positive single-stranded RNA viruses.IMPORTANCE Uncapped viral RNAs often rely on their 5' leader sequences to initiate translation, and the Triticum mosaic virus (TriMV) devotes an astonishing 7% of its genome to directing ribosomes to the correct AUG. Here we uncover a novel mechanism by which a TriMV cis-regulatory element controls cap-independent translation. The upstream region of the functional AUG contains a 16-nt polypyrimidine tract located 11 nt from the initiation site. Based on functional redundancy with similar motifs derived from human picornaviruses, the motif is likely to operate by directing ribosome targeting through base pairing with 18S rRNA. Our results provide the first report of a broad-spectrum mechanism regulating translation initiation for both plant- and animal-hosted picornaviruses.
Keywords: 5′ UTR; IRES; YX-AUG; cap independent; plant virus; potyvirus; translation.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Multiple Cis-acting Polypyrimidine Tract Elements Regulate a Cooperative Mechanism for Triticum Mosaic Virus Internal Ribosomal Entry Site Activity.Front Plant Sci. 2022 Apr 12;13:864832. doi: 10.3389/fpls.2022.864832. eCollection 2022. Front Plant Sci. 2022. PMID: 35498652 Free PMC article.
-
A Unique 5' Translation Element Discovered in Triticum Mosaic Virus.J Virol. 2015 Dec;89(24):12427-40. doi: 10.1128/JVI.02099-15. Epub 2015 Sep 30. J Virol. 2015. PMID: 26423954 Free PMC article.
-
The properties of chimeric picornavirus IRESes show that discrimination between internal translation initiation sites is influenced by the identity of the IRES and not just the context of the AUG codon.RNA. 1999 Jun;5(6):764-78. doi: 10.1017/s1355838299982158. RNA. 1999. PMID: 10376876 Free PMC article.
-
The role of the 5' untranslated regions of Potyviridae in translation.Virus Res. 2015 Aug 3;206:74-81. doi: 10.1016/j.virusres.2015.02.005. Epub 2015 Feb 12. Virus Res. 2015. PMID: 25683508 Review.
-
Hepatitis C Virus Translation Regulation.Int J Mol Sci. 2020 Mar 27;21(7):2328. doi: 10.3390/ijms21072328. Int J Mol Sci. 2020. PMID: 32230899 Free PMC article. Review.
Cited by
-
Multiple Cis-acting Polypyrimidine Tract Elements Regulate a Cooperative Mechanism for Triticum Mosaic Virus Internal Ribosomal Entry Site Activity.Front Plant Sci. 2022 Apr 12;13:864832. doi: 10.3389/fpls.2022.864832. eCollection 2022. Front Plant Sci. 2022. PMID: 35498652 Free PMC article.
-
Conserved RNA secondary structure in Cherry virus A 5'-UTR associated with translation regulation.Virol J. 2022 May 26;19(1):91. doi: 10.1186/s12985-022-01824-z. Virol J. 2022. PMID: 35619168 Free PMC article.
-
RNA elements and their biotechnological applications in plants.New Phytol. 2025 Sep;247(6):2517-2537. doi: 10.1111/nph.70400. Epub 2025 Jul 27. New Phytol. 2025. PMID: 40717389 Free PMC article. Review.
-
Turnip mosaic virus NIb weakens the function of eukaryotic translation initiation factor 6 facilitating viral infection in Nicotiana benthamiana.Mol Plant Pathol. 2024 Feb;25(2):e13434. doi: 10.1111/mpp.13434. Mol Plant Pathol. 2024. PMID: 38388027 Free PMC article.
-
Non-Canonical Translation Initiation Mechanisms Employed by Eukaryotic Viral mRNAs.Biochemistry (Mosc). 2021 Sep;86(9):1060-1094. doi: 10.1134/S0006297921090042. Biochemistry (Mosc). 2021. PMID: 34565312 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
